Articles
- Page Path
- HOME > Endocrinol Metab > Volume 38(5); 2023 > Article
-
Review ArticleDiabetes, obesity and metabolism The Emerging Importance of Mitochondria in White Adipocytes: Neither Last nor Least
 Keypoint
Keypoint
The review underscores the crucial role of mitochondria in controlling white adipose tissue remodeling and energy balance, emphasizing their metabolic versatility, which facilitates tissue adaptability to metabolic signals. Mitochondria influence adipose homeostasis at the cellular level, affecting thermogenic brown and beige adipocytes as well as energy-storing white adipocytes. A comprehensive understanding of mitochondrial regulation in white adipocytes can pave the way for targeted metabolic health strategies. -
Juan Cai1
 , Fenfen Wang2, Mengle Shao1
, Fenfen Wang2, Mengle Shao1 -
Endocrinology and Metabolism 2023;38(5):493-503.
DOI: https://doi.org/10.3803/EnM.2023.1813
Published online: October 10, 2023
1CAS Key Laboratory of Molecular Virology and Immunology, The Center for Microbes, Development and Health, Shanghai Institute of Immunity and Infection, Chinese Academy of Sciences, Shanghai, China
2Department of Anesthesiology, Critical Care and Pain Medicine, Center for Perioperative Medicine, McGovern Medical School, UT Health Science Center at Houston, Houston, TX, USA
- Corresponding authors: Fenfen Wang Department of Anesthesiology, Critical Care and Pain Medicine, Center for Perioperative Medicine, McGovern Medical School, UT Health Science Center at Houston, 6431 Fannin Street, Houston, TX, USA Tel: +1-713-486-0120, Fax: +1-713-500-8922, E-mail: Fenfen.Wang@uth.tmc.edu
- Mengle Shao CAS Key Laboratory of Molecular Virology and Immunology, The Center for Microbes, Development and Health, Shanghai Institute of Immunity and Infection, Chinese Academy of Sciences, 320 Yueyang Road, Shanghai, China Tel: +86-21-5492-3042, Fax: +86-21-5492-3044, E-mail: mlshao@siii.cas.cn
Copyright © 2023 Korean Endocrine Society
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/4.0/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
- 1,975 Views
- 95 Download
- ABSTRACT
- INTRODUCTION
- ROLE OF MITOCHONDRIA IN THERMOGENIC ADIPOCYTES
- ROLE OF MITOCHONDRIA IN WHITE ADIPOCYTE FUNCTION
- WHITE ADIPOCYTE MITOCHONDRIAL HOMEOSTASIS AND METABOLIC HEALTH
- MITOCHONDRIAL REGULATION OF WHITE ADIPOCYTE FORMATION
- MITOCHONDRIAL ROLE IN DETERMINING WHITE ADIPOCYTE IDENTITY
- CONCLUSIONS
- Article information
- References
ABSTRACT
- The growing recognition of mitochondria’s crucial role in the regulation of white adipose tissue remodeling and energy balance underscores its significance. The marked metabolic diversity of mitochondria provides the molecular and cellular foundation for enabling adipose tissue plasticity in response to various metabolic cues. Effective control of mitochondrial function at the cellular level, not only in thermogenic brown and beige adipocytes but also in energy-storing white adipocytes, exerts a profound influence on adipose homeostasis. Furthermore, mitochondria play a pivotal role in intercellular communication within adipose tissue via production of metabolites with signaling properties. A more comprehensive understanding of mitochondrial regulation within white adipocytes will empower the development of targeted and efficacious strategies to enhance adipose function, leading to advancements in overall metabolic health.
- Mitochondria, often referred to as the “powerhouses of the cell,” are remarkable organelles essential for cellular energy production and a host of other vital functions [1-4]. Present in the cytoplasm of eukaryotic cells, these double-membrane-bound structures are derived from ancient bacteria that were engulfed by early eukaryotic cells in a symbiotic relationship. Their primary and most renowned role lies in generating adenosine triphosphate (ATP) through oxidative phosphorylation, a process that efficiently converts nutrients, such as glucose and fatty acids, into usable energy. This metabolic pathway occurs within the inner mitochondrial membrane and is fueled by the electron transport chain and the citric acid cycle. However, mitochondria’s significance extends beyond energy production; they are also involved in regulating cellular metabolism, controlling calcium homeostasis, participating in apoptosis (programmed cell death), and contributing to various signaling pathways [5-9]. Understanding the intricacies of mitochondria and their functions is crucial for comprehending cellular physiology, energy metabolism, and their broader implications for human health and disease.
- White adipocytes are a crucial component of human body’s metabolic machinery. As specialized cells primarily found in adipose tissue, these adipocytes play a central role in energy regulation and storage [10-12]. Their primary function revolves around the storage of triglycerides, which are synthesized from excess dietary fats and carbohydrates. In times of energy surplus, white adipocytes store triglycerides in the form of lipid droplets, effectively acting as energy reservoirs [13,14]. Conversely, during periods of energy demand, such as fasting or physical activity, these adipocytes release stored triglycerides through lipolysis, providing a vital source of energy for the body’s various physiological processes [15]. Beyond their role in energy storage, white adipocytes also secrete a variety of hormones and adipokines that impact systemic metabolism, inflammation, and appetite regulation [16,17].
- Thermogenic brown and beige adipocytes possess a high number of mitochondria, guaranteeing their capacity to uphold distinct bioenergetic processes and effectively transform stored energy into heat [18-20]. Meanwhile, a growing body of evidence is substantiating the critical role of mitochondria in white adipocytes, despite the presence of fewer mitochondria in these cells [21,22].
- Here, we review the current knowledge about the role of mitochondria in white adipocytes, focusing on the mitochondrial regulation of adipocyte formation and function. We also discuss the emerging evidence linking mitochondrial signaling metabolites and white adipocyte identity.
INTRODUCTION
- Mitochondria play a pivotal role in the metabolic dynamics of both brown and beige adipocytes, specialized fat cells that hold significant implications for energy homeostasis and metabolic health [23,24]. Brown adipocytes, prominent in rodents and human infants but also present in human adults, are characterized by a high number of mitochondria and an abundance of iron-rich proteins, giving them their distinctive brown color [25-27]. These mitochondria contain a unique mitochondrial protein called uncoupling protein 1 (UCP1), which enables the dissipation of proton gradients generated during oxidative phosphorylation (OXPHOS) [28,29]. This uncoupling mechanism facilitates the generation of heat rather than ATP, contributing to the thermogenic capacity of brown adipocytes. As a result, brown adipose tissue acts as a natural thermal regulator, aiding in maintaining body temperature and increasing energy expenditure.
- In recent years, beige adipocytes have garnered attention for their capacity to transform into a “brown-like” phenotype in response to various stimuli, such as cold exposure or certain hormonal cues [18,20]. These cells emerge within white adipose tissue (WAT) depots and share similarities with brown adipocytes, including the presence of multilocular lipid droplets and increased mitochondrial content. The mitochondria in beige adipocytes also express UCP1, albeit to a lesser extent than classical brown adipocytes, contributing to their thermogenic capability [20,29]. Notably, creatine, a naturally occurring compound primarily known for its role in cellular energy metabolism, has also garnered attention for its potential involvement in thermogenesis [30]. While traditionally associated with enhancing muscular performance by replenishing ATP, recent research has indicated that creatine may impact thermogenesis through its interactions with mitochondria [31,32]. Studies have suggested that creatine supplementation could enhance mitochondrial bioenergetics and heat production [31,32]. In both brown and beige adipocytes, the central role of mitochondria in thermogenesis and metabolic regulation underscores their importance in maintaining overall energy balance and metabolic health.
ROLE OF MITOCHONDRIA IN THERMOGENIC ADIPOCYTES
- White adipocytes, despite having a lower abundance of mitochondria compared to other types of adipocytes, rely on functional mitochondria as crucial guardians to maintain health and regulate various aspects of adipocyte biology. At a homeostatic level, mitochondria play essential roles in ensuring cellular survival, controlling adipocyte differentiation, maintaining glucose and lipid homeostasis, and orchestrating metabolism of branched-chain amino acids (BCAAs). Additionally, functional mitochondria in white adipocytes significantly contribute to glucose homeostasis and adipokine secretion, and are involved in the browning process, which transforms white adipocytes into thermogenic brown-like adipocytes.
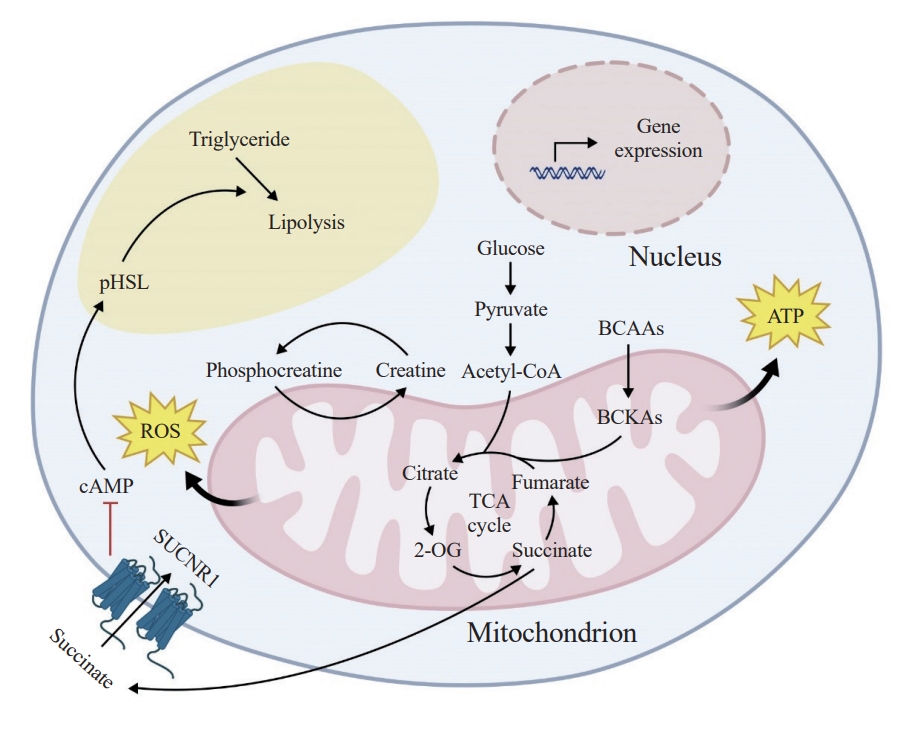
- Like other mitochondria-containing cells, white adipocytes heavily depend on ATP produced by mitochondria to sustain essential metabolic processes such as triglyceride synthesis, gluconeogenesis, and fatty acid re-esterification for survival [33]. This tight relationship is attributed to the mitochondrial energy center, serving as both the hub for cellular energy metabolism and the exclusive site for essential enzymes involved in these anabolic processes [34-36]. For instance, mitochondria play crucial roles in lipogenesis and lipolysis in white adipocytes. This organelle produces critical metabolic intermediates, like citrate, which is essential for the lipogenic process to function optimally [21]. Additionally, mitochondria provide the required environment for lipogenesis and lipolysis, particularly accommodating medium-chain fatty acids [37]. ATP production by mitochondria governs the efficiency of these metabolic pathways. Furthermore, mitochondria actively participate in phospholipid synthesis, further highlighting their multifaceted role in lipid metabolism [38].
- Leucine, isoleucine, and valine are BCAAs and are catabolized only in mitochondria. Given the important influence of adipocytes on BCAA metabolism, mice with depletion of mitochondrial branched-chain amino acid aminotransferase 2 (Bcat2) exhibit an intriguing metabolic profile despite consuming more food. Specifically, these mice show elevated plasma levels of BCAAs and a remarkable reduction in adiposity and body weight. This reduction is accompanied by an increase in energy expenditure, significant improvement in glucose tolerance and insulin sensitivity, and notable protection against diet-induced obesity [39].
- Emerging research emphasizes the pivotal role of mitochondria in governing glucose homeostasis within white adipocytes, particularly through their influence on insulin actions. Notably, when mitochondrial respiratory inhibitors or uncoupling reagents are administered, there is a significant reduction in insulin-stimulated glucose uptake in adipocytes [40].
- Additionally, in rats fed a high-fat diet (HFD), mitochondrial biogenesis and mitochondrial DNA (mtDNA) copy numbers are downregulated in WAT, leading to elevated glucose levels and diminished glucose uptake by white adipocytes [41].
- Mitochondria also play a significant role in adipokine synthesis and secretion, with mitochondrial biogenesis being closely linked to this process. Stimulation of mitochondrial biogenesis through treatments like endothelial nitric oxide synthase activation or overexpression of nuclear respiratory factor-1 (Nrf1) can enhance the synthesis of adiponectin, a critical adipokine [42,43]. Interestingly, adiponectin overexpression reciprocally leads to an increase in mitochondrial density in white adipocytes, creating a regulatory loop between mitochondria and adipokine synthesis [44].
- With the advancement of sequencing technologies, a few studies have investigated the heterogeneity of white adipocytes in mice and humans using single-nucleus RNA sequencing (snRNA-seq) [45-47]. Furthermore, despite limited evidence, it is suggested that the heterogeneity of white adipocytes may be determined by the activity of mitochondria, such as a subpopulation of white adipocytes in mice displaying higher expression of genes associated with mitochondrial activity [45]. Moreover, a sn-RNA-seq study on human white adipocytes revealed two distinct clusters: one with higher gene expression related to mitochondrial capacity and enhanced mitochondrial function, and the other with lower gene expression related to mitochondrial function and reduced mitochondrial activity, respectively [47].
- The mitochondrial unfolded protein response (UPRmt) has a protective effect on white adipocytes, as it serves as a mechanism for mitochondria to manage stress and maintain homeostasis [48,49]. Several studies have demonstrated that UPRmt also plays a crucial role in fat metabolism and protects against obesity. In a cohort study, UPRmt-related genes, including C/EBP homologous protein (Chop), heat shock protein 60 (Hsp60), caseinolytic peptidase (ClpP), and HtrA serine peptidase 2 (Htra2), are significantly reduced in higher-weight identical twins [50]. Moreover, several genes of the UPRmt-related sirtuins (Sirts) family, namely Sirt1/2/3, are involved in the regulation of adipose tissue metabolism [51-56]. It is worth noting that not all UPRmt-related genes are beneficial to the organism, such as ClpP and Park2 (the PARKIN-encoding gene) [57,58]. Deletion of these two factors has been found to result in a beneficial metabolic phenotype.
ROLE OF MITOCHONDRIA IN WHITE ADIPOCYTE FUNCTION
- While the above evidence implies an important role of mitochondrial homeostasis in maintenance of white adipocyte function and health, mitochondrial homeostasis is often disturbed by stress conditions, such as aging, obesity, and other metabolic challenges. When mitochondria turn dysfunctional, they can have adverse effects on white adipocytes and the metabolic homeostasis of the organism. A number of studies have demonstrated that dysfunctional mitochondria in white adipocytes can lead to abnormal lipid metabolism, inflammation, impaired glucose metabolism, dysregulated adipokine secretion, and apoptosis. For example, adipocyte-specific overexpression of amyloidbeta precursor protein (App) has been linked to white adipocyte mitochondrial dysfunction. This results in the inhibition of catecholamine-stimulated lipolysis, leading to significant white adipocyte hypertrophy, increased inflammation, and fibrosis in WAT. These mice also show insulin resistance, glucose intolerance, and sensitivity to obesity [59]. Another critical player in white adipocyte mitochondrial function is the mitochondrial dicarboxylate carrier (mDIC), mainly expressed in WAT and encoded by the Slc25a10 gene. mDIC transfers succinate from the mitochondrial matrix to the cell membrane, where it interacts with the succinate receptor (SUCNR1), blocking the production of cyclic adenosine monophosphate (cAMP) and subsequently suppressing the hormone-sensitive lipase pathway, leading to decreased lipolysis. Dysfunctional mDIC-mediated succinate transport in white adipocytes during obesity results in the release of fatty acids from WAT to the liver, causing adipocyte dysfunction and liver lipotoxicity, thereby impacting the development of nonalcoholic fatty liver disease and systemic insulin resistance [60].
- Mitochondrial dysfunction may have complex effects on the system’s metabolism, such as transcription factor A, mitochondrial (TFAM). Knocking out Tfam specifically in adipocytes leads to mitochondrial dysfunction, decreases weight gain induced by age or diet, impairs glucose tolerance, lowers energy expenditure, promotes inflammation and apoptosis in WAT, and reduces circulating adiponectin and leptin levels. These mice are protected from obesity but are sensitive to hepatosteatosis, hypertension, and cardiac dysfunction [61]. When ferritin mitochondrial (Ftmt), which is a mitochondrial matrix protein that chelates iron, is overexpressed specifically in adipocytes, mice are subjected to dietary challenges, resulting in leaner body weights. However, these mice exhibit dysfunction of adipose metabolism evidenced by glucose intolerance, reduced adiponectin levels, increased damage from reactive oxygen species (ROS), elevated levels of growth/differentiation factor 15 and fibroblast growth factor 21, and decreased circulating and intracellular adiponectin levels. Interestingly, despite the severe adiposity dysfunction in the transgenic mice, a noteworthy outcome is observed—they display significant β-cell proliferation, suggesting a beneficial mitochondria-induced adipose-pancreatic inter-organ signaling axis [62].
- Moreover, mitochondrial dysfunction induced by the deficiency of genes such as CR6-interacting factor 1 (Crif1), FUN14 domain-containing protein 1 (Fundc1), and NADH dehydrogenase (ubiquinone) iron-sulfur protein 4 (Ndufs4) has been associated with increased inflammatory cell infiltration, enhanced inflammatory gene programs and insulin resistance/glucose intolerance [63-65]. Additionally, thymidine kinase 2 (Tk2)-deficiency dramatically reduces circulating levels of leptin and resistin [66]. White adipocytes exposed to mitochondrial dysfunction induced by the administration of oligomycin A and antimycin A and by knockdown of mitochondrial transcription factor A (mtTFA) exhibit a dose-dependent downregulation of adiponectin expression and impaired glucose homeostasis [40]. Iron metabolism also plays an essential role in mitochondrial function, and the manipulation of iron-related genes can lead to mitochondrial dysfunction and abnormal changes in white adipocytes, affecting systemic energy homeostasis. For example, overexpression of Asn-Glu-Glu-Thr (mitoNEET), an outer mitochondrial membrane protein and iron-sulfur cluster transfer protein, causes mitochondrial dysfunction, resulting in increased adipose tissue mass due to enhanced lipid uptake and storage. These mice also display enhanced insulin sensitivity and higher expression of adiponectin [67]. Furthermore, the response of these mice to an HFD challenge is dynamic, with short-term exposure leading to enhanced browning signature in subcutaneous WAT and limited expansion, but with prolonged exposure causing diminished browning and rapid WAT expansion and weight gain [68]. Studies on other iron-related genes, ferritin heavy chain (Fth), and Ftmt, have also provided evidence for the crucial role of mitochondrial iron in WAT functionality [62,69].
- The evidence obtained from rodent models is exciting; however, data from human studies are even more valuable regarding translational implications. Numerous population studies have provided compelling evidence for the significant association between adipocyte mitochondrial function and obesity. For instance, lower body weight and higher mitochondrial respiration rate and mitochondrial number in WAT show strong correlations [70]. Comparative studies involving identical twins have revealed that the obese twins had significantly lower mitochondrial mtDNA than their non-obese counterparts, despite both having the same mtDNA sequence [71]. A research study conducted in Japanese and Italian populations has demonstrated a significant association between mtDNA variation at position 15497 guanine/adenine (Mt15497G→A) and lipid metabolism as well as obesity-related factors. These factors include body weight, body mass index, waist circumference, hip circumference, and intra-abdominal fat [72,73]. Another study has investigated the status of subcutaneous WAT mitochondrial OXPHOS in four groups: non-obese individuals, non-obese individuals with type 2 diabetes mellitus (T2DM), nondiabetic obese individuals, and obese individuals with T2DM. In both non-obese and obese T2DM subjects, mitochondrial transmembrane potential, inorganic phosphate utilization, and electron transport chain activity are significantly reduced compared to the corresponding nonobese individuals without T2DM. Notably, the mitochondrial index, a measure of mitochondrial function, is significantly higher in lean individuals with T2DM than in lean individuals without diabetes. Furthermore, in people with obesity and T2DM, respiratory chain activities (specifically, complex I, I to III, II to III) and phosphorylation capacity of white adipocyte mitochondria are markedly lower than in obese individuals without T2DM [74]. Collectively, these findings indicate that obesity itself can have a detrimental impact on white adipocyte mitochondrial function, leading to various metabolic disturbances associated with obesity.
WHITE ADIPOCYTE MITOCHONDRIAL HOMEOSTASIS AND METABOLIC HEALTH
- In addition to the crucial role of mitochondria in white adipocyte function, a growing number of studies suggest that mitochondrial regulation is pivotal for the cell lineage fate of adipocyte progenitors and the formation of mature white adipocytes.
- Given the above-mentioned mitochondrial role in BCAA metabolism, a substantial increase in leucine catabolism and expression of enzymes involved in the BCAA catabolic pathway is observed with increasing mitochondrial numbers during adipogenesis [75,76]. Adipogenesis is also concomitantly accompanied by heightened mitochondrial biogenesis, activity, and the generation of ROS [21,33]. Consequently, the differentiation process reprograms the energy status from glycolytic to oxidative metabolism, thus acquiring and sustaining a mature adipocyte phenotype [77]. Consequently, any malfunction in mitochondrial metabolism or disruption in ROS production, induced either through genetic manipulation of mitochondrial constituents or employment of pharmacological inhibitors, is sufficient to perturb adipogenesis, evidenced in studies using cultured preadipocyte cell lines and the stromal vascular fraction of adipose tissues [78-80].
- With recent discoveries of adipose tissue-resident adipose progenitor cells (APCs), more precise and targeted evidence pertaining to mitochondrial regulation of their lineage potential has come to light [81]. Of note, unbiased quantitative proteomic analysis of murine APC populations has revealed significant depot- and sex-dependent differences in the expression of mitochondrial proteins and regulators of lipid metabolism [82,83].
- Functionally, impaired mitochondrial activity (achieved by a physiologically relevant reduction via ectopic expression of the mitoNEET protein, the above-mentioned outer mitochondrial membrane protein governing oxidative capacity) exerts a profound influence on the behavior of platelet-derived growth factor receptor (PDGFR) β+ adipose progenitors, driving them to adopt a pro-inflammatory phenotype at the cost of their adipogenic capacity in adult mice [84].
MITOCHONDRIAL REGULATION OF WHITE ADIPOCYTE FORMATION
- White adipocytes possess remarkable phenotypical plasticity and dynamism, allowing them to undergo a transformative process known as browning or beiging when exposed to certain conditions such as cold temperatures, activation of β-adrenergic receptors, exercise or diet [85-89]. During this process, white adipocytes adopt characteristics similar to brown adipocytes, resulting in the induction of beige adipocytes. Evidently, a crucial aspect of this browning/beiging process involves increasing the number of mitochondria and enhancing mitochondrial biogenesis. This mitochondrial enhancement is a key factor in facilitating the transition of white adipocytes towards a more thermogenic and metabolically active phenotype, thus promoting energy expenditure and metabolic health [90].
- Mitochondrial double-membraned structures encompass a large variety of chemical reactions that occur within an organism to sustain life, including the breakdown of nutrients and the synthesis of essential molecules [2,91]. Metabolites, the small molecules involved in these metabolic processes, serve as building blocks, energy sources, and signaling molecules that regulate various cellular activities [92,93]. Emerging evidence is showing that mitochondrial signaling metabolites act as intricate messengers, transmitting information about cellular status, environmental cues, and metabolic processes, ensuring the efficient cell fate determination of different cells, including adipocyte lineage cells. By influencing enzymatic activities, gene expression, and signaling pathways, signal metabolites orchestrate a wide array of physiological responses, spanning from adaptation to stress to the coordination of growth and differentiation [5]. Succinate, a key metabolite in the tricarboxylic acid cycle, has recently garnered significant attention for its intriguing role in the modulation of WAT physiology [60,94]. Emerging research has unveiled a novel connection between succinate and the conversion of white adipocytes to beige adipocytes, a process known as white-to-beige adipocyte conversion. It is proposed that succinate is involved in the activation of pathways related to beige adipogenesis and thermogenesis [95].
- Our recent work highlighted that mitoprotease Lon protease 1 (LONP1)-dependent metabolic enzyme turnover controls whiteto-beige adipocyte conversion via fine-tuning intracellular succinate concentration [96]. We observed an increase in mitochondrial protein turnover, coupled with dynamic adjustments in the levels of the pivotal mitoprotease LONP1, in response to thermogenic stimulation. We uncovered the selective degradation of succinate dehydrogenase B (SDHB), a crucial element of mitochondrial complex II, by LONP1. Following thermogenic stimulation, this LONP1-mediated SDHB degradation led to an elevation in intracellular succinate levels. The resultant rise in the succinate to α-ketoglutarate ratio fostered enhanced histone methylation on thermogenic genes, thereby promoting the shift from white-to-beige adipocytes. Notably, elevating LONP1 expression induced heightened intracellular succinate levels, effectively rejuvenating beige cell conversion and adaptive thermogenesis in aged adipocytes and mice.
MITOCHONDRIAL ROLE IN DETERMINING WHITE ADIPOCYTE IDENTITY
- In recent years, extensive research has been directed towards unraveling the intricacies of mitochondrial regulation in maintaining metabolic homeostasis within adipocytes, which includes the emerging body of evidence that highlights the notable importance of mitochondria within white adipocytes (Fig. 1). These advancements have greatly enhanced our comprehension of the pivotal roles played by mitochondria in ensuring the overall health of adipose tissue. The orchestration of mitochondrial activity is indispensable in shaping the functionality of mature adipocytes, while also exerting a profound influence on the microenvironment within WAT.
- The exploration of therapeutic applicability of these findings remains at the forefront in the field of WAT mitochondrial research. A comprehensive analysis of the temporal progression of each WAT mitochondrial dysfunction model documented in prior research becomes instrumental in ascertaining the optimal window for initiating therapeutic interventions aimed at remedying the observed mitochondrial aberrations. Understanding the complex interplay between mitochondrial function and adipocyte physiology as well as its implication in the development of pathological WAT microenvironment is another forefront of adipocyte mitochondrial biology.
- In addition, mitochondrial metabolism is emerging as a key hub of signal metabolite production, linking mitochondrial functional normality to active regulation of various biological processes, such as cell fate determination, beyond traditional views of mitochondrial metabolism that is passively adapted to meet energy demands of the cell. Further understanding of the intricate interplay between signaling metabolites (e.g., succinate), adipocyte biology, and energy metabolism holds promise for the development of innovative therapeutic strategies aimed at enhancing energy expenditure and combating obesity.
CONCLUSIONS
-
CONFLICTS OF INTEREST
No potential conflict of interest relevant to this article was reported.
Article information
-
Acknowledgements
- This work was supported by the National Natural Science Foundation of China (82170891), Shanghai Municipal Science and Technology Major Project (2019SHZDZX02), Shanghai Municipal Science and Technology Project (22140903200), and Shanghai Pujiang Program (21PJ1414600) to Mengle Shao. Juan Cai was also supported by China Postdoctoral Science Foundation (2022M723264), Shanghai Postdoctoral Excellence Program (2022644), Special Research Assistant Funding Project of Chinese Academy of Sciences.
- 1. McBride HM, Neuspiel M, Wasiak S. Mitochondria: more than just a powerhouse. Curr Biol 2006;16:R551–60.ArticlePubMed
- 2. Friedman JR, Nunnari J. Mitochondrial form and function. Nature 2014;505:335–43.ArticlePubMedPMCPDF
- 3. Picard M, Shirihai OS. Mitochondrial signal transduction. Cell Metab 2022;34:1620–53.ArticlePubMedPMC
- 4. Monzel AS, Enriquez JA, Picard M. Multifaceted mitochondria: moving mitochondrial science beyond function and dysfunction. Nat Metab 2023;5:546–62.ArticlePubMedPMCPDF
- 5. Spinelli JB, Haigis MC. The multifaceted contributions of mitochondria to cellular metabolism. Nat Cell Biol 2018;20:745–54.ArticlePubMedPMCPDF
- 6. Giorgi C, Marchi S, Pinton P. The machineries, regulation and cellular functions of mitochondrial calcium. Nat Rev Mol Cell Biol 2018;19:713–30.ArticlePubMedPDF
- 7. Garbincius JF, Elrod JW. Mitochondrial calcium exchange in physiology and disease. Physiol Rev 2022;102:893–992.ArticlePubMedPMC
- 8. Bock FJ, Tait SW. Mitochondria as multifaceted regulators of cell death. Nat Rev Mol Cell Biol 2020;21:85–100.ArticlePubMedPDF
- 9. Tan JX, Finkel T. Mitochondria as intracellular signaling platforms in health and disease. J Cell Biol 2020;219:e202002179.ArticlePubMedPMCPDF
- 10. Sethi JK, Vidal-Puig AJ. Thematic review series: adipocyte biology: adipose tissue function and plasticity orchestrate nutritional adaptation. J Lipid Res 2007;48:1253–62.ArticlePubMedPMC
- 11. Morigny P, Boucher J, Arner P, Langin D. Lipid and glucose metabolism in white adipocytes: pathways, dysfunction and therapeutics. Nat Rev Endocrinol 2021;17:276–95.ArticlePubMedPDF
- 12. Rosen ED, Spiegelman BM. What we talk about when we talk about fat. Cell 2014;156:20–44.ArticlePubMedPMC
- 13. Sakers A, De Siqueira MK, Seale P, Villanueva CJ. Adiposetissue plasticity in health and disease. Cell 2022;185:419–46.ArticlePubMed
- 14. Zechner R, Zimmermann R, Eichmann TO, Kohlwein SD, Haemmerle G, Lass A, et al. FAT SIGNALS: lipases and lipolysis in lipid metabolism and signaling. Cell Metab 2012;15:279–91.ArticlePubMedPMC
- 15. Lafontan M, Langin D. Lipolysis and lipid mobilization in human adipose tissue. Prog Lipid Res 2009;48:275–97.ArticlePubMed
- 16. Fasshauer M, Bluher M. Adipokines in health and disease. Trends Pharmacol Sci 2015;36:461–70.ArticlePubMed
- 17. Funcke JB, Scherer PE. Beyond adiponectin and leptin: adipose tissue-derived mediators of inter-organ communication. J Lipid Res 2019;60:1648–84.ArticlePubMedPMC
- 18. Wang W, Seale P. Control of brown and beige fat development. Nat Rev Mol Cell Biol 2016;17:691–702.ArticlePubMedPMCPDF
- 19. Shamsi F, Wang CH, Tseng YH. The evolving view of thermogenic adipocytes: ontogeny, niche and function. Nat Rev Endocrinol 2021;17:726–44.ArticlePubMedPMCPDF
- 20. Cohen P, Kajimura S. The cellular and functional complexity of thermogenic fat. Nat Rev Mol Cell Biol 2021;22:393–409.ArticlePubMedPMCPDF
- 21. Heinonen S, Jokinen R, Rissanen A, Pietilainen KH. White adipose tissue mitochondrial metabolism in health and in obesity. Obes Rev 2020;21:e12958.ArticlePubMedPDF
- 22. Zhu Q, An YA, Scherer PE. Mitochondrial regulation and white adipose tissue homeostasis. Trends Cell Biol 2022;32:351–64.ArticlePubMedPMC
- 23. Sidossis L, Kajimura S. Brown and beige fat in humans: thermogenic adipocytes that control energy and glucose homeostasis. J Clin Invest 2015;125:478–86.ArticlePubMedPMC
- 24. Aquilano K, Zhou B, Brestoff JR, Lettieri-Barbato D. Multifaceted mitochondrial quality control in brown adipose tissue. Trends Cell Biol 2023;33:517–29.ArticlePubMed
- 25. Cypess AM, Lehman S, Williams G, Tal I, Rodman D, Goldfine AB, et al. Identification and importance of brown adipose tissue in adult humans. N Engl J Med 2009;360:1509–17.ArticlePubMedPMC
- 26. van Marken Lichtenbelt WD, Vanhommerig JW, Smulders NM, Drossaerts JM, Kemerink GJ, Bouvy ND, et al. Coldactivated brown adipose tissue in healthy men. N Engl J Med 2009;360:1500–8.ArticlePubMed
- 27. Virtanen KA, Lidell ME, Orava J, Heglind M, Westergren R, Niemi T, et al. Functional brown adipose tissue in healthy adults. N Engl J Med 2009;360:1518–25.ArticlePubMed
- 28. Fedorenko A, Lishko PV, Kirichok Y. Mechanism of fattyacid-dependent UCP1 uncoupling in brown fat mitochondria. Cell 2012;151:400–13.ArticlePubMedPMC
- 29. Chouchani ET, Kazak L, Spiegelman BM. New advances in adaptive thermogenesis: UCP1 and beyond. Cell Metab 2019;29:27–37.ArticlePubMed
- 30. Kazak L, Chouchani ET, Jedrychowski MP, Erickson BK, Shinoda K, Cohen P, et al. A creatine-driven substrate cycle enhances energy expenditure and thermogenesis in beige fat. Cell 2015;163:643–55.ArticlePubMedPMC
- 31. Sun Y, Rahbani JF, Jedrychowski MP, Riley CL, Vidoni S, Bogoslavski D, et al. Mitochondrial TNAP controls thermogenesis by hydrolysis of phosphocreatine. Nature 2021;593:580–5.ArticlePubMedPMCPDF
- 32. Rahbani JF, Roesler A, Hussain MF, Samborska B, Dykstra CB, Tsai L, et al. Creatine kinase B controls futile creatine cycling in thermogenic fat. Nature 2021;590:480–5.ArticlePubMedPMCPDF
- 33. Lee JH, Park A, Oh KJ, Lee SC, Kim WK, Bae KH. The role of adipose tissue mitochondria: regulation of mitochondrial function for the treatment of metabolic diseases. Int J Mol Sci 2019;20:4924.ArticlePubMedPMC
- 34. Pagliarini DJ, Rutter J. Hallmarks of a new era in mitochondrial biochemistry. Genes Dev 2013;27:2615–27.ArticlePubMedPMC
- 35. Wiedemann N, Pfanner N. Mitochondrial machineries for protein import and assembly. Annu Rev Biochem 2017;86:685–714.ArticlePubMed
- 36. Ernster L, Schatz G. Mitochondria: a historical review. J Cell Biol 1981;91(3 Pt 2):227s–55s.ArticlePubMedPMCPDF
- 37. De Pauw A, Tejerina S, Raes M, Keijer J, Arnould T. Mitochondrial (dys)function in adipocyte (de)differentiation and systemic metabolic alterations. Am J Pathol 2009;175:927–39.ArticlePubMedPMC
- 38. Vance JE. Phospholipid synthesis and transport in mammalian cells. Traffic 2015;16:1–18.ArticlePubMed
- 39. She P, Reid TM, Bronson SK, Vary TC, Hajnal A, Lynch CJ, et al. Disruption of BCATm in mice leads to increased energy expenditure associated with the activation of a futile protein turnover cycle. Cell Metab 2007;6:181–94.ArticlePubMedPMC
- 40. Wang CH, Wang CC, Huang HC, Wei YH. Mitochondrial dysfunction leads to impairment of insulin sensitivity and adiponectin secretion in adipocytes. FEBS J 2013;280:1039–50.ArticlePubMedPDF
- 41. Sutherland LN, Capozzi LC, Turchinsky NJ, Bell RC, Wright DC. Time course of high-fat diet-induced reductions in adipose tissue mitochondrial proteins: potential mechanisms and the relationship to glucose intolerance. Am J Physiol Endocrinol Metab 2008;295:E1076–83.ArticlePubMed
- 42. Koh EH, Kim M, Ranjan KC, Kim HS, Park HS, Oh KS, et al. eNOS plays a major role in adiponectin synthesis in adipocytes. Am J Physiol Endocrinol Metab 2010;298:E846–53.ArticlePubMed
- 43. Koh EH, Park JY, Park HS, Jeon MJ, Ryu JW, Kim M, et al. Essential role of mitochondrial function in adiponectin synthesis in adipocytes. Diabetes 2007;56:2973–81.ArticlePubMedPDF
- 44. Asterholm IW, Scherer PE. Enhanced metabolic flexibility associated with elevated adiponectin levels. Am J Pathol 2010;176:1364–76.ArticlePubMedPMC
- 45. Rajbhandari P, Arneson D, Hart SK, Ahn IS, Diamante G, Santos LC, et al. Single cell analysis reveals immune celladipocyte crosstalk regulating the transcription of thermogenic adipocytes. Elife 2019;8:e49501.ArticlePubMedPMCPDF
- 46. Sarvari AK, Van Hauwaert EL, Markussen LK, Gammelmark E, Marcher AB, Ebbesen MF, et al. Plasticity of epididymal adipose tissue in response to diet-induced obesity at single-nucleus resolution. Cell Metab 2021;33:437–53.ArticlePubMed
- 47. Whytock KL, Sun Y, Divoux A, Yu G, Smith SR, Walsh MJ, et al. Single cell full-length transcriptome of human subcutaneous adipose tissue reveals unique and heterogeneous cell populations. iScience 2022;25:104772.ArticlePubMedPMC
- 48. Yoneda T, Benedetti C, Urano F, Clark SG, Harding HP, Ron D. Compartment-specific perturbation of protein handling activates genes encoding mitochondrial chaperones. J Cell Sci 2004;117(Pt 18):4055–66.ArticlePubMedPDF
- 49. Zhao Q, Wang J, Levichkin IV, Stasinopoulos S, Ryan MT, Hoogenraad NJ. A mitochondrial specific stress response in mammalian cells. EMBO J 2002;21:4411–9.ArticlePubMedPMC
- 50. Jukarainen S, Heinonen S, Ramo JT, Rinnankoski-Tuikka R, Rappou E, Tummers M, et al. Obesity is associated with low NAD(+)/SIRT pathway expression in adipose tissue of BMIdiscordant monozygotic twins. J Clin Endocrinol Metab 2016;101:275–83.ArticlePubMed
- 51. Milne JC, Lambert PD, Schenk S, Carney DP, Smith JJ, Gagne DJ, et al. Small molecule activators of SIRT1 as therapeutics for the treatment of type 2 diabetes. Nature 2007;450:712–6.ArticlePubMedPMCPDF
- 52. Lagouge M, Argmann C, Gerhart-Hines Z, Meziane H, Lerin C, Daussin F, et al. Resveratrol improves mitochondrial function and protects against metabolic disease by activating SIRT1 and PGC-1alpha. Cell 2006;127:1109–22.ArticlePubMed
- 53. Picard F, Kurtev M, Chung N, Topark-Ngarm A, Senawong T, Machado De Oliveira R, et al. Sirt1 promotes fat mobilization in white adipocytes by repressing PPAR-gamma. Nature 2004;429:771–6.ArticlePubMedPMCPDF
- 54. Bordone L, Cohen D, Robinson A, Motta MC, van Veen E, Czopik A, et al. SIRT1 transgenic mice show phenotypes resembling calorie restriction. Aging Cell 2007;6:759–67.ArticlePubMed
- 55. Jing E, Gesta S, Kahn CR. SIRT2 regulates adipocyte differentiation through FoxO1 acetylation/deacetylation. Cell Metab 2007;6:105–14.ArticlePubMedPMC
- 56. Shi T, Wang F, Stieren E, Tong Q. SIRT3, a mitochondrial sirtuin deacetylase, regulates mitochondrial function and thermogenesis in brown adipocytes. J Biol Chem 2005;280:13560–7.ArticlePubMed
- 57. Bhaskaran S, Pharaoh G, Ranjit R, Murphy A, Matsuzaki S, Nair BC, et al. Loss of mitochondrial protease ClpP protects mice from diet-induced obesity and insulin resistance. EMBO Rep 2018;19:e45009.ArticlePubMedPMCPDF
- 58. Moore TM, Cheng L, Wolf DM, Ngo J, Segawa M, Zhu X, et al. Parkin regulates adiposity by coordinating mitophagy with mitochondrial biogenesis in white adipocytes. Nat Commun 2022;13:6661.ArticlePubMedPMCPDF
- 59. An YA, Crewe C, Asterholm IW, Sun K, Chen S, Zhang F, et al. Dysregulation of amyloid precursor protein impairs adipose tissue mitochondrial function and promotes obesity. Nat Metab 2019;1:1243–57.ArticlePubMedPMCPDF
- 60. An YA, Chen S, Deng Y, Wang ZV, Funcke JB, Shah M, et al. The mitochondrial dicarboxylate carrier prevents hepatic lipotoxicity by inhibiting white adipocyte lipolysis. J Hepatol 2021;75:387–99.ArticlePubMedPMC
- 61. Vernochet C, Damilano F, Mourier A, Bezy O, Mori MA, Smyth G, et al. Adipose tissue mitochondrial dysfunction triggers a lipodystrophic syndrome with insulin resistance, hepatosteatosis, and cardiovascular complications. FASEB J 2014;28:4408–19.ArticlePubMedPMCPDF
- 62. Kusminski CM, Ghaben AL, Morley TS, Samms RJ, Adams AC, An Y, et al. A novel model of diabetic complications: adipocyte mitochondrial dysfunction triggers massive β-cell hyperplasia. Diabetes 2020;69:313–30.ArticlePubMedPMCPDF
- 63. Ryu MJ, Kim SJ, Kim YK, Choi MJ, Tadi S, Lee MH, et al. Crif1 deficiency reduces adipose OXPHOS capacity and triggers inflammation and insulin resistance in mice. PLoS Genet 2013;9:e1003356.ArticlePubMedPMC
- 64. Wu H, Wang Y, Li W, Chen H, Du L, Liu D, et al. Deficiency of mitophagy receptor FUNDC1 impairs mitochondrial quality and aggravates dietary-induced obesity and metabolic syndrome. Autophagy 2019;15:1882–98.ArticlePubMedPMC
- 65. Choi KM, Ryan KK, Yoon JC. Adipose mitochondrial complex I deficiency modulates inflammation and glucose homeostasis in a sex-dependent manner. Endocrinology 2022;163:bqac018.ArticlePubMedPMCPDF
- 66. Villarroya J, Dorado B, Vila MR, Garcia-Arumi E, Domingo P, Giralt M, et al. Thymidine kinase 2 deficiency-induced mitochondrial DNA depletion causes abnormal development of adipose tissues and adipokine levels in mice. PLoS One 2011;6:e29691.ArticlePubMedPMC
- 67. Kusminski CM, Holland WL, Sun K, Park J, Spurgin SB, Lin Y, et al. MitoNEET-driven alterations in adipocyte mitochondrial activity reveal a crucial adaptive process that preserves insulin sensitivity in obesity. Nat Med 2012;18:1539–49.ArticlePubMedPMCPDF
- 68. Kusminski CM, Park J, Scherer PE. MitoNEET-mediated effects on browning of white adipose tissue. Nat Commun 2014;5:3962.ArticlePubMedPMCPDF
- 69. Blankenhaus B, Braza F, Martins R, Bastos-Amador P, Gonzalez-Garcia I, Carlos AR, et al. Ferritin regulates organismal energy balance and thermogenesis. Mol Metab 2019;24:64–79.ArticlePubMedPMC
- 70. Ling Y, Carayol J, Galusca B, Canto C, Montaurier C, Matone A, et al. Persistent low body weight in humans is associated with higher mitochondrial activity in white adipose tissue. Am J Clin Nutr 2019;110:605–16.ArticlePubMedPMCPDF
- 71. Mustelin L, Pietilainen KH, Rissanen A, Sovijarvi AR, Piirila P, Naukkarinen J, et al. Acquired obesity and poor physical fitness impair expression of genes of mitochondrial oxidative phosphorylation in monozygotic twins discordant for obesity. Am J Physiol Endocrinol Metab 2008;295:E148–54.ArticlePubMedPMC
- 72. Liguori R, Mazzaccara C, Pasanisi F, Buono P, Oriani G, Finelli C, et al. The mtDNA 15497 G/A polymorphism in cytochrome b in severe obese subjects from Southern Italy. Nutr Metab Cardiovasc Dis 2006;16:466–70.ArticlePubMed
- 73. Okura T, Koda M, Ando F, Niino N, Tanaka M, Shimokata H. Association of the mitochondrial DNA 15497G/A polymorphism with obesity in a middle-aged and elderly Japanese population. Hum Genet 2003;113:432–6.ArticlePubMedPDF
- 74. Chattopadhyay M, Guhathakurta I, Behera P, Ranjan KR, Khanna M, Mukhopadhyay S, et al. Mitochondrial bioenergetics is not impaired in nonobese subjects with type 2 diabetes mellitus. Metabolism 2011;60:1702–10.ArticlePubMed
- 75. Frerman FE, Sabran JL, Taylor JL, Grossberg SE. Leucine catabolism during the differentiation of 3T3-L1 cells. Expression of a mitochondrial enzyme system. J Biol Chem 1983;258:7087–93.ArticlePubMed
- 76. Kitsy A, Carney S, Vivar JC, Knight MS, Pointer MA, Gwathmey JK, et al. Effects of leucine supplementation and serum withdrawal on branched-chain amino acid pathway gene and protein expression in mouse adipocytes. PLoS One 2014;9:e102615.ArticlePubMedPMC
- 77. Kladnicka I, Cedikova M, Kripnerova M, Dvorakova J, Kohoutova M, Tuma Z, et al. Mitochondrial respiration of adipocytes differentiating from human mesenchymal stem cells derived from adipose tissue. Physiol Res 2019;68(Suppl 3):S287–96.PubMed
- 78. Kusminski CM, Scherer PE. Mitochondrial dysfunction in white adipose tissue. Trends Endocrinol Metab 2012;23:435–43.ArticlePubMedPMC
- 79. Tormos KV, Anso E, Hamanaka RB, Eisenbart J, Joseph J, Kalyanaraman B, et al. Mitochondrial complex III ROS regulate adipocyte differentiation. Cell Metab 2011;14:537–44.ArticlePubMedPMC
- 80. Fujiwara M, Tian L, Le PT, DeMambro VE, Becker KA, Rosen CJ, et al. The mitophagy receptor Bcl-2-like protein 13 stimulates adipogenesis by regulating mitochondrial oxidative phosphorylation and apoptosis in mice. J Biol Chem 2019;294:12683–94.ArticlePubMedPMC
- 81. Maniyadath B, Zhang Q, Gupta RK, Mandrup S. Adipose tissue at single-cell resolution. Cell Metab 2023;35:386–413.ArticlePubMedPMC
- 82. Shan B, Barker CS, Shao M, Zhang Q, Gupta RK, Wu Y. Multilayered omics reveal sex- and depot-dependent adipose progenitor cell heterogeneity. Cell Metab 2022;34:783–99.ArticlePubMedPMC
- 83. Shao M, Hepler C, Zhang Q, Shan B, Vishvanath L, Henry GH, et al. Pathologic HIF1α signaling drives adipose progenitor dysfunction in obesity. Cell Stem Cell 2021;28:685–701.ArticlePubMedPMC
- 84. Joffin N, Paschoal VA, Gliniak CM, Crewe C, Elnwasany A, Szweda LI, et al. Mitochondrial metabolism is a key regulator of the fibro-inflammatory and adipogenic stromal subpopulations in white adipose tissue. Cell Stem Cell 2021;28:702–17.ArticlePubMedPMC
- 85. Shore AM, Karamitri A, Kemp P, Speakman JR, Graham NS, Lomax MA. Cold-induced changes in gene expression in brown adipose tissue, white adipose tissue and liver. PLoS One 2013;8:e68933.ArticlePubMedPMC
- 86. de Jong JM, Wouters RT, Boulet N, Cannon B, Nedergaard J, Petrovic N. The β3-adrenergic receptor is dispensable for browning of adipose tissues. Am J Physiol Endocrinol Metab 2017;312:E508–18.ArticlePubMed
- 87. Bostrom P, Wu J, Jedrychowski MP, Korde A, Ye L, Lo JC, et al. A PGC1-α-dependent myokine that drives brown-fatlike development of white fat and thermogenesis. Nature 2012;481:463–8.ArticlePubMedPMCPDF
- 88. Scheel AK, Espelage L, Chadt A. Many ways to Rome: exercise, cold exposure and diet: do they all affect BAT activation and WAT browning in the same manner? Int J Mol Sci 2022;23:4759.ArticlePubMedPMC
- 89. Wang F, Xiao F, Du L, Niu Y, Yin H, Zhou Z, et al. Activation of GCN2 in macrophages promotes white adipose tissue browning and lipolysis under leucine deprivation. FASEB J 2021;35:e21652.ArticlePubMedPDF
- 90. Lu X. Maintaining mitochondria in beige adipose tissue. Adipocyte 2019;8:77–82.ArticlePubMedPMCPDF
- 91. Giacomello M, Pyakurel A, Glytsou C, Scorrano L. The cell biology of mitochondrial membrane dynamics. Nat Rev Mol Cell Biol 2020;21:204–24.ArticlePubMedPDF
- 92. Martinez-Reyes I, Chandel NS. Mitochondrial TCA cycle metabolites control physiology and disease. Nat Commun 2020;11:102.ArticlePubMedPMCPDF
- 93. Baker SA, Rutter J. Metabolites as signalling molecules. Nat Rev Mol Cell Biol 2023;24:355–74.ArticlePubMedPDF
- 94. Villanueva-Carmona T, Cedo L, Madeira A, Ceperuelo-Mallafre V, Rodriguez-Pena MM, Nunez-Roa C, et al. SUCNR1 signaling in adipocytes controls energy metabolism by modulating circadian clock and leptin expression. Cell Metab 2023;35:601–19.ArticlePubMed
- 95. Mills EL, Pierce KA, Jedrychowski MP, Garrity R, Winther S, Vidoni S, et al. Accumulation of succinate controls activation of adipose tissue thermogenesis. Nature 2018;560:102–6.ArticlePubMedPMCPDF
- 96. Fu T, Sun W, Xue J, Zhou Z, Wang W, Guo Q, et al. Proteolytic rewiring of mitochondria by LONP1 directs cell identity switching of adipocytes. Nat Cell Biol 2023;25:848–64.ArticlePubMedPDF
 PubReader
PubReader ePub Link
ePub Link Cite
Cite