Articles
- Page Path
- HOME > Endocrinol Metab > Volume 38(5); 2023 > Article
-
Original ArticleDiabetes, obesity and metabolism Protective Effects of Melatonin in High-Fat Diet-Induced Hepatic Steatosis via Decreased Intestinal Lipid Absorption and Hepatic Cholesterol Synthesis
 Keypoint
Keypoint
The study explored melatonin's preventive effects on obesity and fatty liver development under a high-fat diet in mice and HepG2 cells. Melatonin reduced hepatic cholesterol synthesis, as indicated by decreased expression of SREBP-2, HMGCR, and LDLR. It also reduced intestinal carbohydrate and lipid absorption, evidenced by decreased expression of SGLT1, GLUT2, GLUT5, and NPC1L1. The study concludes that melatonin's protective effects against weight gain and liver steatosis under a high-fat diet are due to reduced caloric absorption and cholesterol synthesis. -
Hyungjune Ku1
 , Yeonji Kim1, Alvin Lyle Kim2, Garam Lee3, Youngsik Choi1, Bukyung Kim1
, Yeonji Kim1, Alvin Lyle Kim2, Garam Lee3, Youngsik Choi1, Bukyung Kim1 -
Endocrinology and Metabolism 2023;38(5):557-567.
DOI: https://doi.org/10.3803/EnM.2023.1672
Published online: September 1, 2023
1Department of Internal Medicine, Kosin University College of Medicine, Busan, Korea
2Department of Surgery, Kosin University College of Medicine, Busan, Korea
3Department of Food Science and Nutrition, Pusan National University, Busan, Korea
- Corresponding author: Bukyung Kim Division of Endocrinology and Metabolism, Department of Internal Medicine, Kosin University College of Medicine, 262 Gamcheon-ro, Seo-gu, Busan 49267, Korea Tel: +82-51-990-6102, Fax: +82-51-248-5686, E-mail: 79kyung@daum.net
Copyright © 2023 Korean Endocrine Society
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/4.0/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
- 1,991 Views
- 94 Download
- 1 Crossref
ABSTRACT
-
Background
- The preventative effect of melatonin on the development of obesity and the progression of fatty liver under a high-fat diet (HFD) has been well elucidated through previous studies. We investigated the mechanism behind this effect regarding cholesterol biosynthesis and regulation of cholesterol levels.
-
Methods
- Mice were divided into three groups: normal chow diet (NCD); HFD; and HFD and melatonin administration group (HFD+M). We assessed the serum lipid profile, mRNA expression levels of proteins involved in cholesterol synthesis and reabsorption in the liver and nutrient transporters in the intestines, and cytokine levels. Additionally, an in vitro experiment using HepG2 cells was performed.
-
Results
- Expression of hepatic sterol regulatory element-binding protein 2 (SREBP-2), 3-hydroxy-3-methylglutaryl coenzyme A reductase (HMGCR), and low-density lipoprotein receptor (LDLR) demonstrated that melatonin administration significantly reduces hepatic cholesterol synthesis in mice fed an HFD. Expression of intestinal sodium-glucose transporter 1 (SGLT1), glucose transporter 2 (GLUT2), GLUT5, and Niemann-pick C1-like 1 (NPC1L1) demonstrated that melatonin administration significantly reduces intestinal carbohydrate and lipid absorption in mice fed an HFD. There were no differences in local and circulatory inflammatory cytokine levels among the NCD, HFD, and HFD+M group. HepG2 cells stimulated with palmitate showed reduced levels of SREBP, LDLR, and HMGCR indicating these results are due to the direct mechanistic effect of melatonin on hepatocytes.
-
Conclusion
- Collectively, these data indicate the mechanism behind the protective effects of melatonin from weight gain and liver steatosis under HFD is through a reduction in intestinal caloric absorption and hepatic cholesterol synthesis highlighting its potential in the treatment of obesity and fatty liver disease.
- The worldwide prevalence of obesity has nearly tripled since 1975 to 1 out of 10 adults being obese (body mass index [BMI] ≥30 kg/m2) and 4 out of 10 being overweight (BMI ≥25 kg/m2) as of 2016, according to the World Health Organization [1]. This trend of the global increase in obesity is also present in the adolescent population despite some regional disparities. On the contrary to the high-income Western countries which showed the smallest increase in adolescent obesity since 2000, the highest increase was seen in the east Asian region [2]. This trend is also prominent in the adolescents of South Korea, as well as other east Asian countries, who are exposed to sleep deprivation due to the excessive pressure to study for college entrance exams [3,4]. Such an increase in the prevalence of obesity in the adolescent population will lead to various metabolic disorders in the future which consequently carry a high risk of cardiovascular complications and mortality. Therefore, this will not only be a problem at an individual level but also a factor of serious socioeconomic burden [3].
- Sleep deprivation leads to the state of chronodisruption which is known to increase obesity, risk of diabetes, cardiovascular disease, fatty liver disease, and mortality [5-11]. Previous studies have shown that sleep deprivation leads to obesity and diabetes which could also be achieved by the elimination of melatonin through the removal of the pineal gland [12]. It is also known that reversal of these negative effects could be achieved through melatonin administration in these cases [13].
- Increasing evidence indicates that melatonin deficiency, due to lifestyle changes in modern society is involved in the pathogenesis of the recent worldwide increase in obesity [14,15]. Furthermore, recent discussions are highlighting the potential role of melatonin as a prevention and treatment for metabolic obesity and its complications. Hence, intensive research is being conducted to investigate the various mechanisms of its beneficial effects such as anti-inflammatory effect, insulin sensitivity, and activation of brown adipose tissues [9,16,17].
- Cholesterol synthesis and cholesterol level regulation plays an important role in obesity and its complications [18-21]. Most studies have shown that melatonin inhibits cholesterol synthesis, but it is controversial whether it is 3-hydroxy-3-methylglutaryl coenzyme A reductase (HMGCR) related [22]. In obesity development, the first step is the intestinal absorption of calories. But, only a few studies have investigated whether melatonin is involved in fat absorption [23]. In particular, there have been no studies on nutrient transporters related to lipid and glucose absorption.
- Therefore, in the present study, we hypothesized that the mechanism by which melatonin suppresses obesity is due to the inhibition of cholesterol synthesis in the liver and nutrient absorption in the intestine. We evaluated the inhibitory effects of melatonin in obesity and fatty liver induced by a high-fat diet (HFD) through mRNA expression levels of proteins involved in cholesterol synthesis and reabsorption in the liver and nutrient transporters related to glucose and lipid absorption in the intestines. Additionally, we investigated whether this is a direct mechanistic effect of melatonin on hepatocytes.
INTRODUCTION
- Animal experiments
- Four-week-old male C57BL/6 mice were purchased from Orient Bio (Seongnam, Korea). The mice were housed in a humidity-controlled room maintained at 24℃ under a 12-hour lightdark cycle, with free access to different food and water. All animal experiments were performed according to the Animal Experimentation Ethics Committee of Kosin University (KMAP20-8). The mouse was divided into three groups of five mice each: (1) normal diet group (fed standard rodent chow, contains 5.54% calories from fat, 74% calories from carbohydrates, and 21% calories from protein); (2) HFD group (fed HFD that contained 60% fat, 20% protein, 20% carbohydrate); and (3) melatonin group (fed on HFD and administered melatonin). The melatonin (50 mg/kg body weight) was injected into the intraperitoneal cavity 5 days per week around the afternoon for 17 weeks. Melatonin receptors 1 (MT1) and 2 (MT2) are expressed in the liver and intestine and administration of melatonin increased the content of MT1/MT2 receptor protein in a dose-dependent manner [24,25]. Melatonin at 50 mg/kg dose had previously been shown anti-inflammatory, neuroprotective, and antiobesity effects in rats and mice [26-28].
- Body weight and liver weight evaluation
- Body weight was measured every week in the morning (10:00 AM to 11:00 AM). Liver weight was measured after mouse sacrifice.
- Histological analysis
- The liver was removed from each mouse, fixed 4% paraformaldehyde, paraffin-embedded and sectioned at 4 μm. Hematoxylin and eosin (H&E) staining was according to the standard method. Frozen sections liver, 4 µm-thick cryosections, were stained with Oil red O (Sigma-Aldrich, St. Louis, MO, USA). All sections were observed with a microscope.
- Total cholesterol, apolipoprotein A1, and apolipoprotein B level analysis
- All level measurement was determined in the serum. Total cholesterol level was quantified with QuickDetect total cholesterol (mouse) enzyme-linked immunosorbent assay (ELISA) kit (Bio-Vision, Milpitas, CA, USA). Apolipoprotein A1 (ApoA1) and apolipoprotein B (ApoB) levels were measured with Mouse ApoA1 ELISA kit, mouse ApoB ELISA kit (Abcam, Cambridge, UK). All kit was according to the manufacturer’s instructions.
- Cell culture and treatments
- The human liver cancer cell line HepG2 was cultured in 75-cm2 plastic flasks (Falcon-Becton Dickinson Labwares, Franklin Lakes, NJ, USA) and maintained at 37°C with 5% CO2 in air atmosphere in Dulbecco’s modified essential medium (DMEM) supplemented with 10% (volume/volume) heat-inactivated fetal bovine serum (FBS) and antibiotics (100 U/mL of penicillin, 100 lg/mL of streptomycin). HepG2 cells were treated to 250 or 500 μM palmitate (Sigma-Aldrich) with or without 1.5 mM melatonin conjugated to fatty acid-free bovine serum albumin (BSA) for 24 hours. HepG2 cells exposed with BSA were used as control.
- Reverse transcriptase-polymerase chain reaction
- Total RNA was isolated using TRIzol reagent (Invitrogen, Carlsbad, CA, USA) and reverse transcription was performed with superscript reverse transcriptase (Invitrogen) according to the manufacturer’s instruction. Total RNA (1 μg) was used to prepare cDNA. Expression of HMGCR, low-density lipoprotein receptor (LDLR), sterol regulatory element-binding protein 2 (SREBP-2) at the liver tissue and sodium-glucose transporter 1 (SGLT1), glucose transporter 5 (GLUT5), GLUT2, Niemannpick C1-like 1 (NPC1L1) at the small intestinal epithelium were determined by polymerase chain reaction (PCR). The primers used for amplification were as follows: HMGCR forward, 5´-TGAGATCCGGAGGATCCAAG-3´ and reverse, 5´-ATCCAGCGACTATGAGCGTG-3´, LDLR forward, 5´-TTTGGAGGATGAGAACCGGC-3´ and reverse, 5´-CAGGTACTGGCAACCACCAT -3´, SREBP-2 forward, 5´-CTGGAGACCATGGAGACCCTC-3´ and reverse 5´-CAGGTACTGGCAACCACCAT-3´, SGLT1 forward, 5´-GCCATGGGTGGCTTTGAATG-3´ and reverse, 5´-GAGCAGGGACAGAACGGAAA-3´, GLUT5 forward, 5´-ACCAAGGGAGCCATCAACAAG-3´ and reverse, 5´-GTTATCCTGACTGCTGCCGTA-3´, GLUT2 forward, 5´-ACCGGGATGATTGGCATGTT-3´ and reverse 5´-GGACCTGGCCCAATCTCAAA-3´, NPC1L1 forward, 5´-CCGCGTTCCGAAACCTTGCC-3´ and reverse 5´- AGGATTGCGAAAACACCTGC-3´. Cycling parameters for the three amplifications consisted of an initial DNA denaturation at 94°C for 5 minutes followed by 35 cycles at 94°C for 20 seconds, 55°C for 20 seconds, 72°C for 20 seconds, and final extension at 72°C for 5 minutes. Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) was used as an internal control.
- Quantitative polymerase chain reaction
- Total RNA from mouse small intestine tissue was reverse transcribed using TOPscript cDNA Synthesis kit (Ezynomics Inc., Daejeon, Korea) following the manufactures recommendations. Quantitatve PCR was performed using the QuantStudio3 real-time PCR (Applied Biosystems, Waltham, MA, USA) and TB Green premix Ex Taq II kit according to the manufactures directions. The primers used for amplification were as follows: HMGCR forward, 5´-ATCAGCTGCACCATGCCATC-3´ and reverse, 5´-GTGCAAGCTCCTTGGAGGTC-3´, LDLR forward, 5´-AAGGACACAGCACACAACCA-3´ and reverse, 5´-CAAAGGAAGAAGACGAGGAGCAC-3´, SREBP-2 forward, 5´-CTGACTTCCCTGCTGCAGTG-3´ and reverse 5´- GTTTCCGGTGCCTCCAGAAG-3´, Expression levels for each sample, normalized to GAPDH expression, were calculated as the average of two measurements.
- Immunofluorescence staining
- The HepG2 cells were cultured on collagen-coated 4-well glass chamber slides in DMEM media containing 10% FBS. Cells were fixed in 4% paraformaldehyde/phosphate-buffered saline (PBS) followed by permeabilization in 0.2% Triton X-100/PBS. 3-Hydroxy-3-methylglutaryl coenzyme A (HMGCoA) was obtained from Santa Cruz Biotechnology (Santa Cruz, CA, USA). Appropriate primary antibodies were added to the cells for 1 hour at 37°C. For secondary labeling, cells were incubated with fluorescein-conjugated secondary antibodies (Invitrogen) for 1 hour at 37°C. Nuclei were counterstained using Deep Red (Invitrogen). Fluorescent images were observed and analyzed under fluorescence microscopy (Eclipse 80, Nikon, Tokyo, Japan).
- Western blot analysis
- For Western blots analysis, equivalent amounts of total protein (15 mg) were loaded onto 10% sodium dodecyl sulfate-polyacrylamide gels electrophoresis (SDS/PAGE). The gels were transferred to nitrocellulose membranes using an electro-blotting apparatus (Bio-Rad, Hercules, CA, USA) and reacted with antibodies according to standard methods. The proteins were then transferred to a nitrocellulose membrane using an electroblotting apparatus (Bio-Rad), and the membranes were incubated with each primary antibody (Anti-HMGCoA, LDLR antibodies, Santa Cruz Biotechnology). Blots were incubated with a horseradish peroxidase conjugated anti-mouse secondary antibody (Santa Cruz Biotechnology). The membranes were developed using an enhanced chemiluminescence reaction system and visualized with an Amersham Imager 600 (GE Healthcare, Chicago, IL, USA). β-Actin was used as an internal control to monitor equal protein loading.
- Biochemical analysis
- Plasma and liver protein levels of the pro-inflammatory cytokines interleukin 6 (IL-6), IL-10, and tumor necrosis factor α (TNF-α) were measured using commercial ELISA kits (mouse IL-6 uncoated ELISA, and mouse TNF-α uncoated ELISA, Invitrogen). All experimental steps were performed at room temperature and in duplicate. Samples were quantified by spectrophotometry at 450 nm.
METHODS
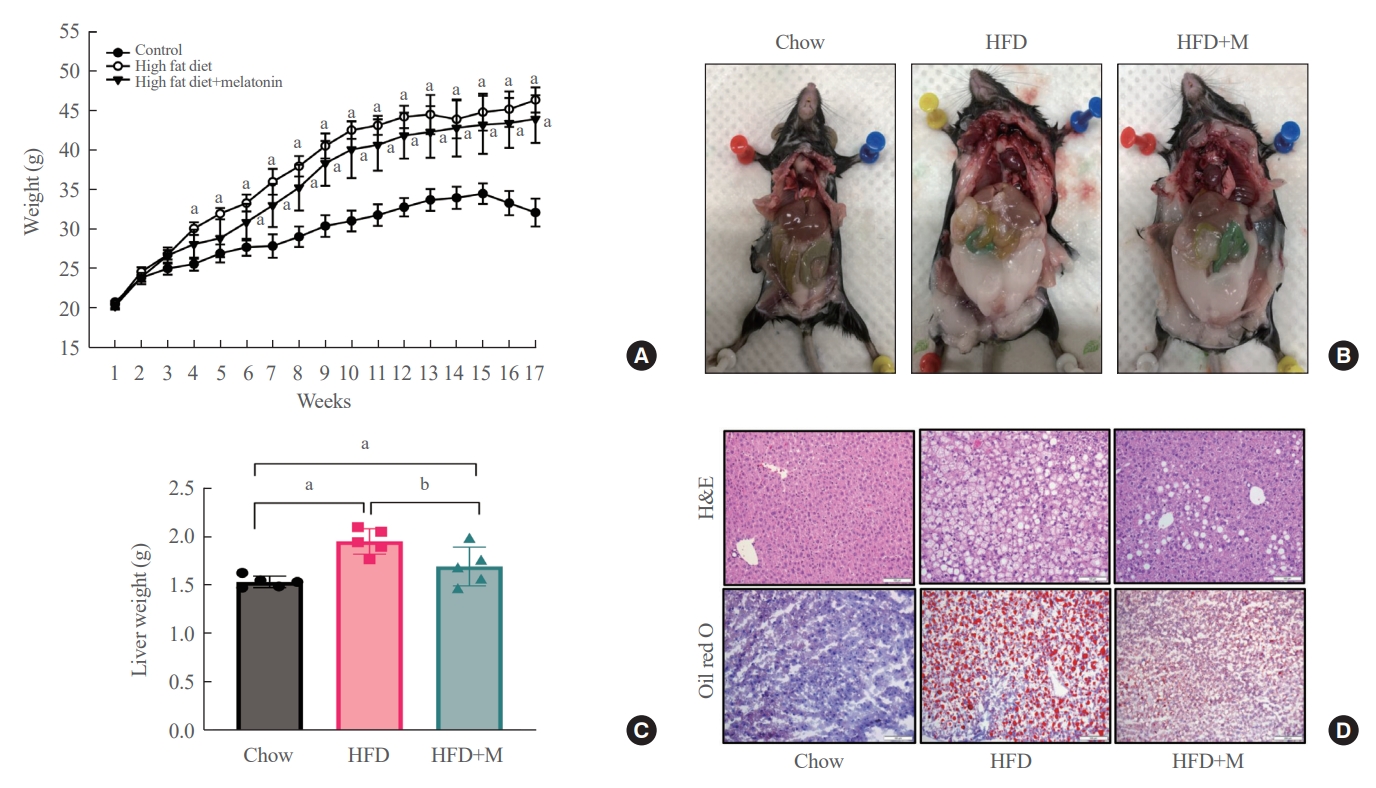
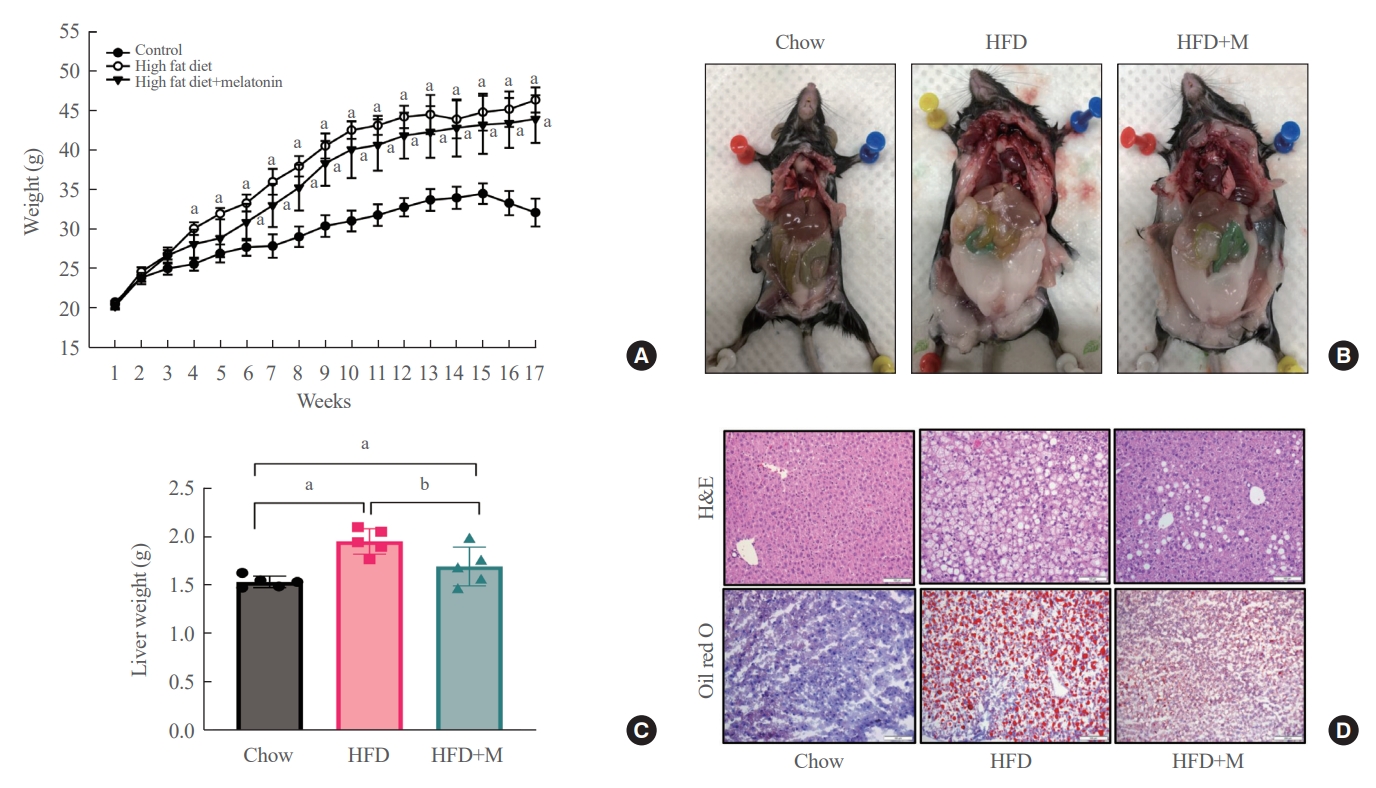
- Melatonin attenuates HFD-induced weight gain, visceral fat accumulation, and liver steatosis
- Weight gain, visceral fat accumulation, and steatosis are major contributors to metabolic obesity and its complications [29]. Therefore, we first examined these factors in mice under a normal chow diet (NCD), HFD, and HFD with melatonin administration (HFD+M) for 17 weeks. As expected, compared to the HFD mice which gained the most weight, reduced weight gain was seen in HFD+M mice while NCD mice gained the least weight (Fig. 1A). The same trend could be observed upon visual inspection of visceral and subcutaneous fat accumulation via gross examination, and extent of lipid accumulation in liver tissues via histological assessments as shown in Fig. 1B, D. In both cases, melatonin decreased fat accumulation in HFD. Liver weights, consistent with the body weight gain, were significantly heavier in HFD mice compared to the NCD mice, and melatonin administration significantly prevented liver weight gain in HFD+M mice (Fig. 1C).
- Melatonin ameliorates HFD-induced elevation of lowd-ensity lipoprotein cholesterol
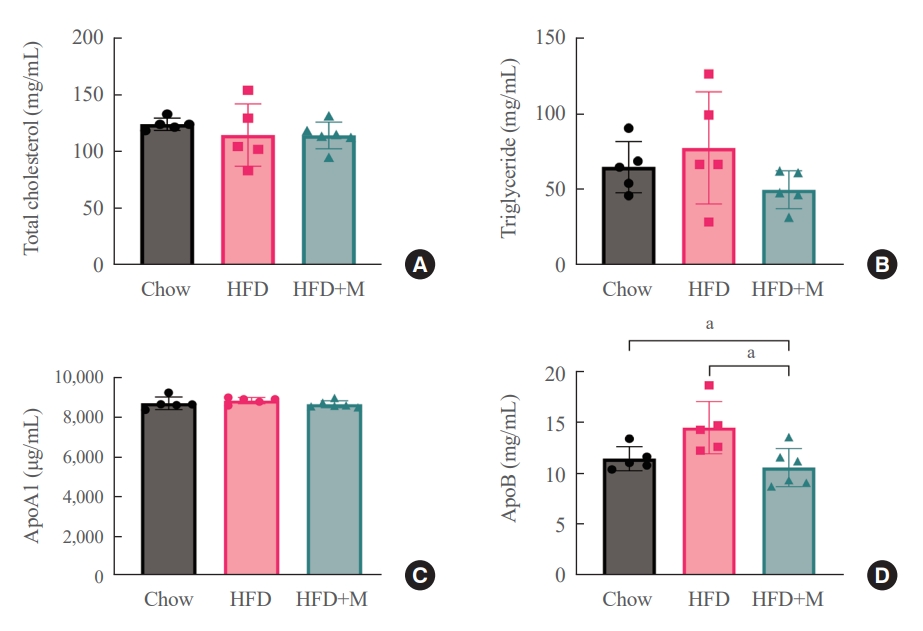
- One of the driving forces for metabolic obesity and its complications, namely hepatic steatosis in HFD is hyperlipidemia [30]. In this aspect, serum lipid profiles were measured at 6 weeks to gain a better understanding of the mechanisms underlying the protective effects of melatonin in HFD. No significant differences were observed among the three groups in terms of total cholesterol (Fig. 2A) and ApoA1 lipoprotein levels (Fig. 2C). However, serum ApoB lipoprotein levels were significantly lower in the HFD+M mice compared to the HFD mice (Fig. 2D), and a similar trend was observed for the serum triglyceride levels as well (Fig. 2B).
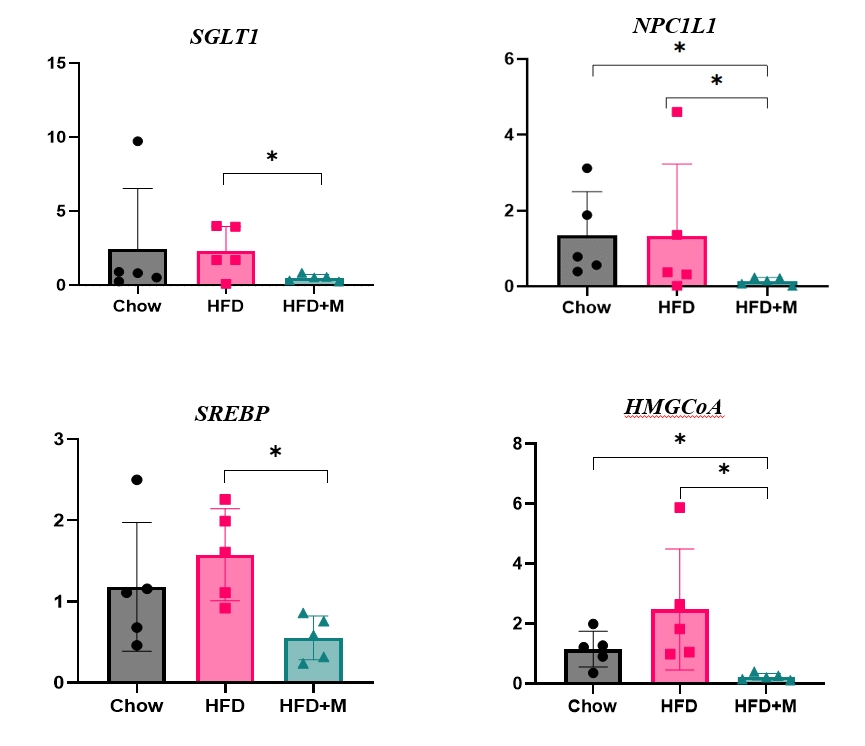
- Protective effects of melatonin from absorption of excess calories via suppression of carbohydrate and lipid transporter expressions in the intestine
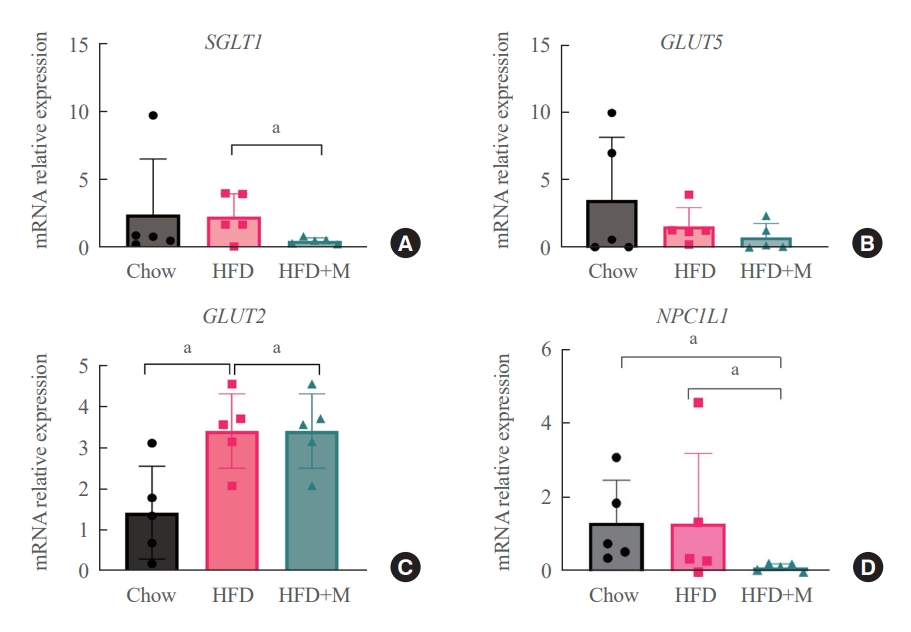
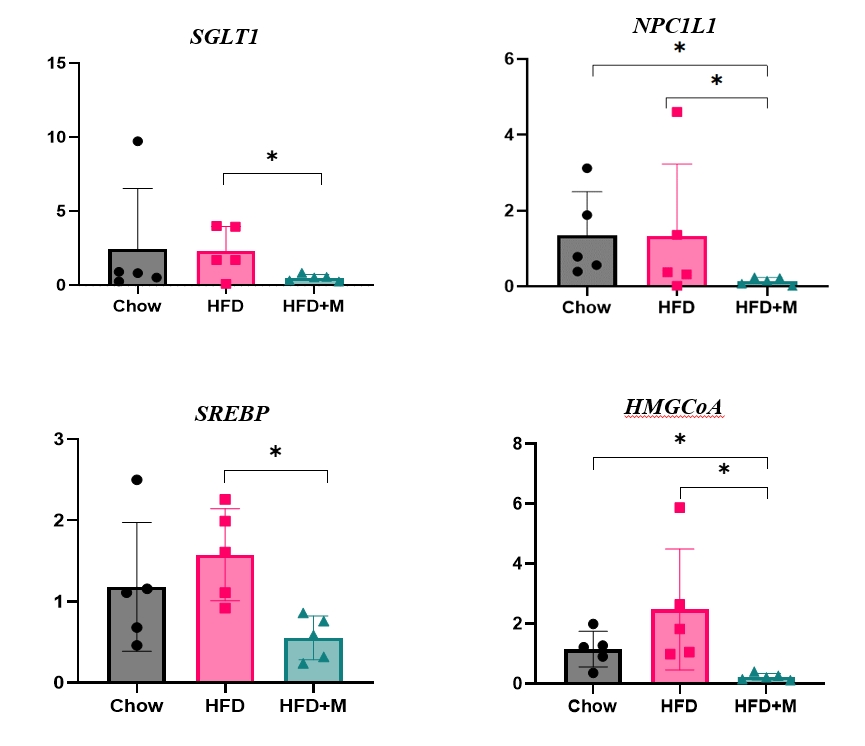
- The fact that melatonin inhibits the production of chylomicrons in the intestines prompted us to investigate its effect on intestinal lipid absorption under HFD. To examine the effect of melatonin on intestinal caloric absorption, small intestinal expressions of various nutrient transporters were assessed. Interestingly, intestinal carbohydrate transporters, SGLT1 at the brush border and GLUT2 at the basolateral membranes were significantly suppressed by melatonin compared to HFD mice (Fig. 3A, C). GLUT5 at the brush border also showed a strong trend to be suppressed by melatonin compared to HFD mice (Fig. 3B). Moreover, NPC1L1, a cholesterol transporter in the liver and the intestines was also significantly suppressed by melatonin compared to HFD and NCD mice as shown in Fig. 3D.
- Melatonin suppresses hepatic cholesterol synthesis
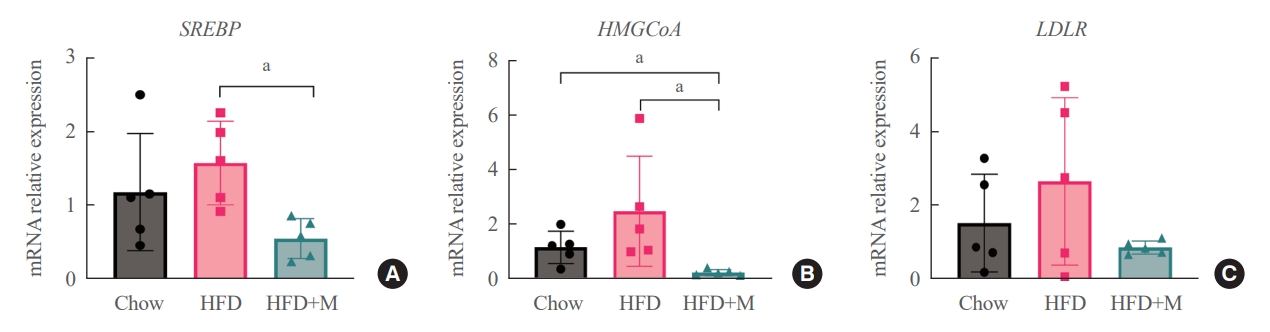
- Given the central role of the liver in the regulation of low-density lipoprotein (LDL) cholesterol levels, we further investigated the effect of melatonin on hepatic cholesterol synthesis. Therefore, we evaluated the mRNA expression levels of genes involved in hepatic cholesterol metabolism. mRNA expression levels of genes that promote cholesterol biosynthesis, SREBP-2, and HMGCR were increased by HFD [18,20]. However, melatonin administration significantly inhibited the expression levels of both genes under HFD (Fig. 4A, B). A similar pattern was seen in the expression levels of LDLR, which promotes hepatic cholesterol reuptake, although not at a statistically significant level (Fig. 4C).
- No differences in circulatory and local inflammatory cytokine levels between groups
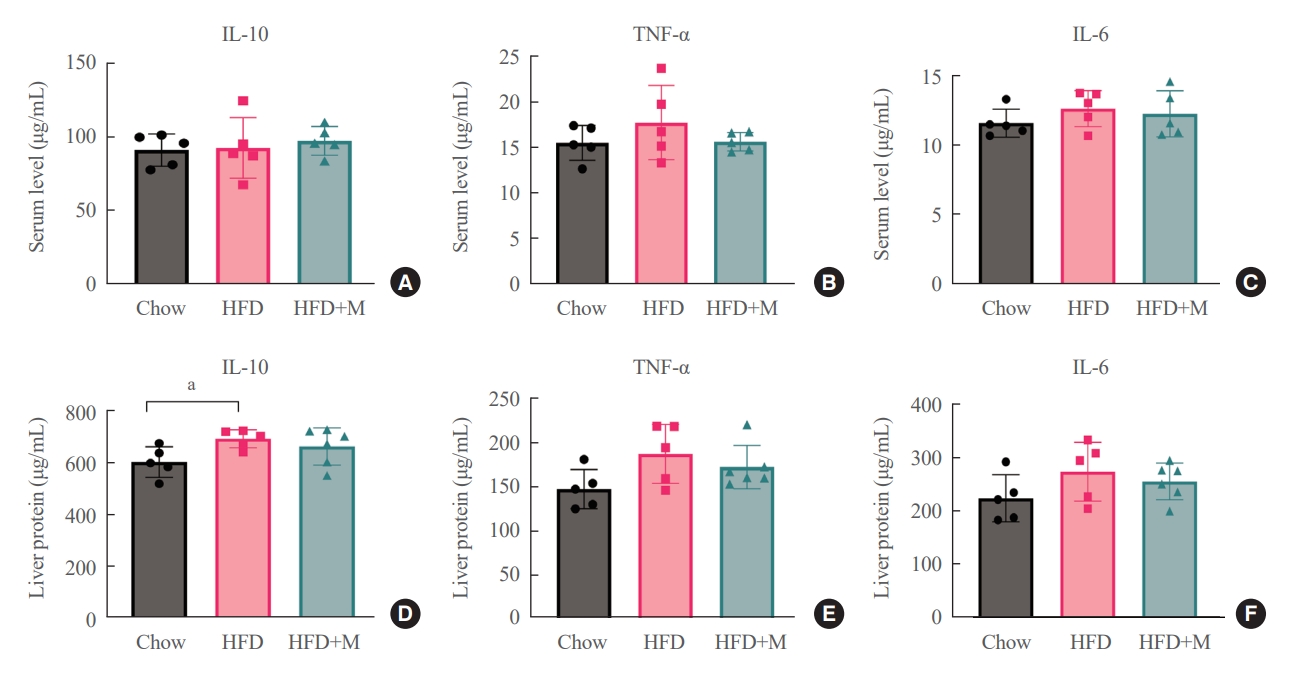
- To determine whether these protective effects of melatonin on the inhibition of hepatic steatosis and obesity are through its direct action on target cells, we measured local and circulatory inflammatory cytokine levels. Measurements of local inflammatory cytokine levels were taken from the liver. As shown in Fig. 5, there were no significant differences in local and circulatory inflammatory cytokine levels between the three groups except for the liver IL-10 level, which was higher in HFD mice compared to NCD mice.
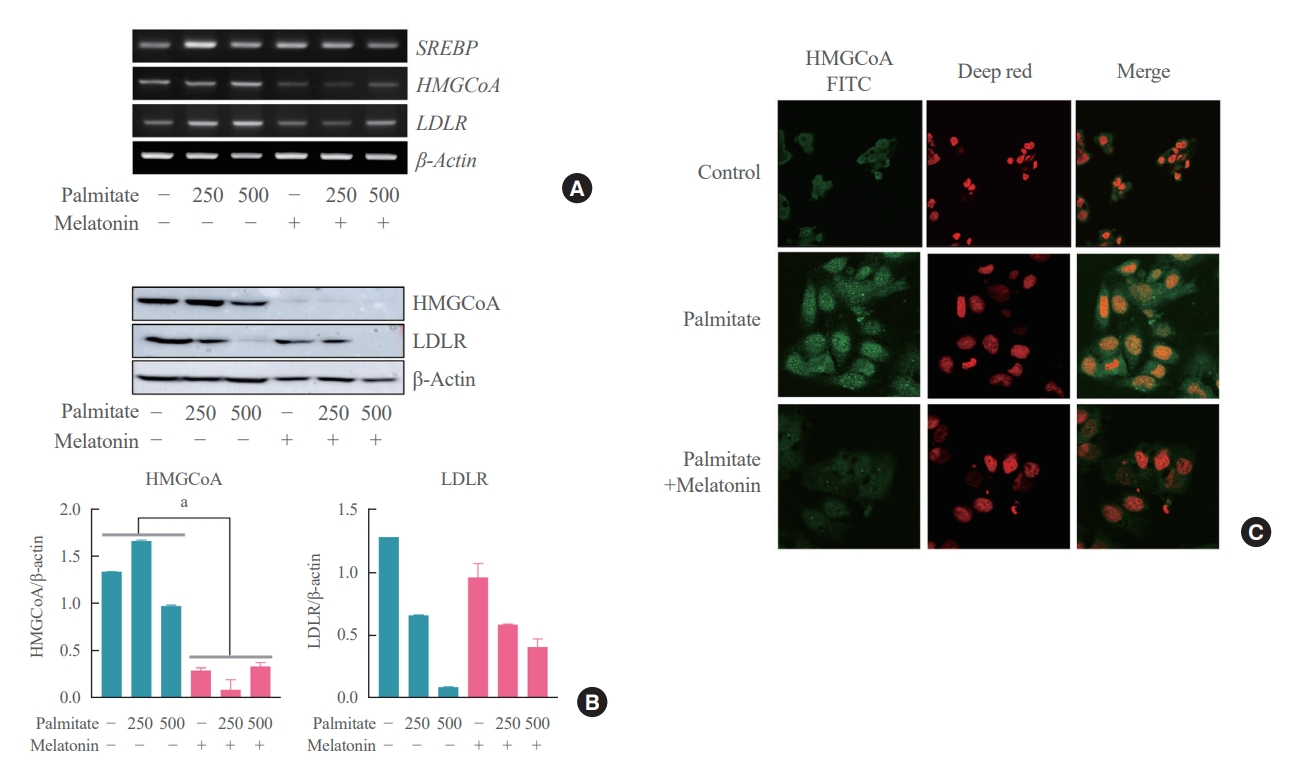
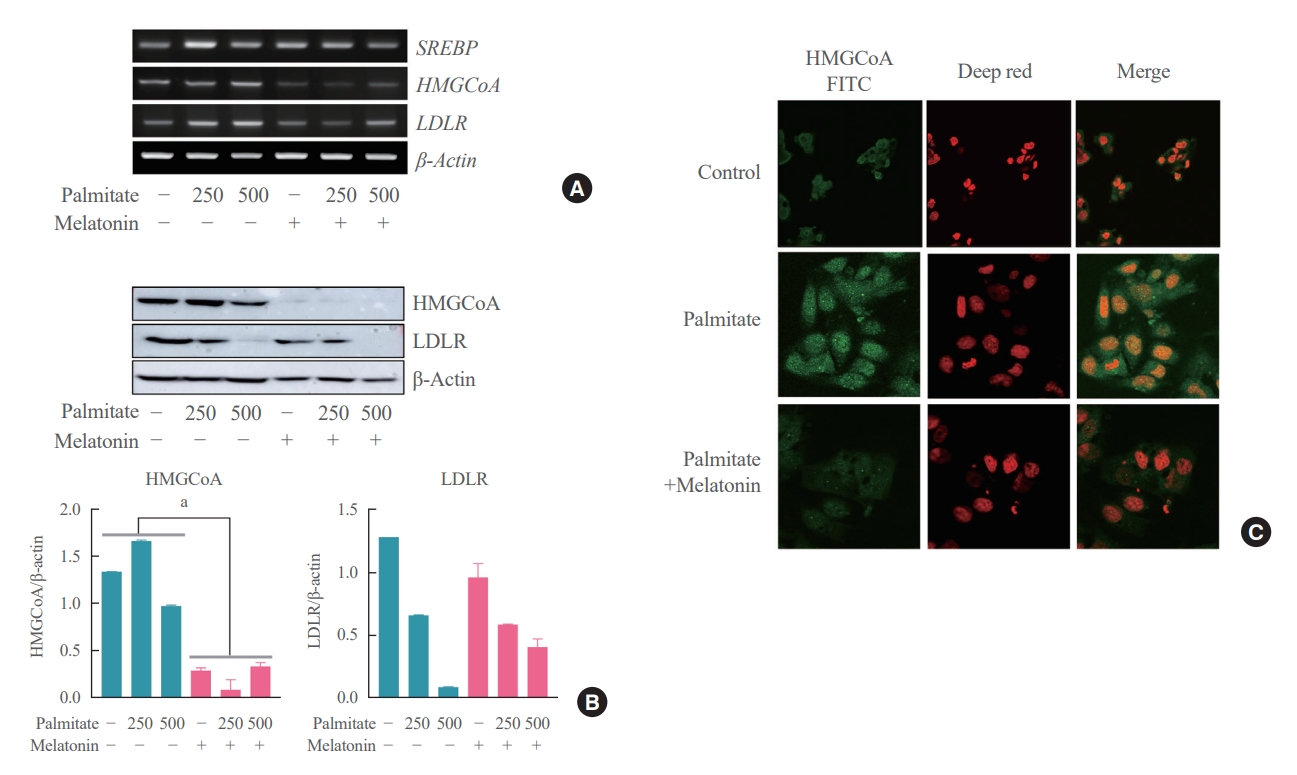
- Melatonin directly inhibits palmitate-induced cholesterol synthesis in hepatocytes
- Lastly, we attempted to address the potential concerns regarding the existence of other indirect pathways underlying the mechanism of action of melatonin on the hepatocytes. All hepatocytes were treated with palmitate for 6 hours, and the melatonin group was concurrently treated with melatonin for 16 hours compared to the control group which was not treated with melatonin. As a result, expression levels of SREBP-2, HMGCR, and LDLR were all decreased in the melatonin group compared to the control group as shown in Fig. 6A. We confidently demonstrated that melatonin directly inhibits cholesterol synthesis in hepatocytes through Western blot and immunofluorescence staining showing the same pattern as shown in Fig. 6B, C.
RESULTS
- The preventative effect of melatonin on the development of obesity and the progression of fatty liver under HFD has been well elucidated through previous studies [31,32]. The majority of these studies have identified the mechanism behind this effect in the aspect of gut microbiota modulation [17], anti-inflammatory, and anti-oxidative properties of melatonin [9,30]. However, cholesterol biosynthesis and regulation of cholesterol level is an integral part of the development of obesity and its complications [18-21] and currently, the mechanistic effect of melatonin in this aspect requires further clarity.
- After 17 weeks, HFD+M mice demonstrated decreased body weight gain and visceral fat accumulation compared to HFD mice (Fig. 1A, B). Also, concurrent administration of melatonin significantly prevented weight gain of the liver compared to HFD alone (Fig. 1C). Additionally, upon histological analysis of the liver tissue using H&E and Oil red O stain, the extent of lipid accumulation was less in the HFD+M mice compared to HFD mice (Fig. 1D). Collectively, these observations indicate the preventative effect of melatonin in HFD-induced weight gain, visceral fat accumulation, and steatosis which is in alignment with previous studies.
- In the current study, we sought to further explore the mechanism underlying this effect in the aspect of serum lipid profiles. In terms of total cholesterol and ApoA1 lipoprotein, no differences were seen among the three groups (Fig. 2A, C). Interestingly, the level of ApoB lipoprotein, which is a constituent of LDL cholesterol, was significantly decreased in the HFD+M mice compared to HFD mice (Fig. 2D). A similar pattern was seen for the serum triglyceride levels although not at a statistically significant level (Fig. 2B). Dietary lipid digest into cholesterol, triglyceride, and free fatty acid, and after absorption, they turn into chylomicrons to transport to the liver. The liver excretes very low density lipoprotein (VLDL) to the systemic blood, and after using triglyceride for energy fuel whole body’s LDL remains. These are ApoB-contained apolipoproteins. Whereas ApoA1 is related to high-density lipoprotein cholesterol [33,34]. Therefore, these results indicate the inhibitory effect of melatonin in production of chylomicrons in the intestines and LDL cholesterol in the liver.
- Dietary cholesterol is absorbed in the intestinal epithelium, goes through esterification, and enters the circulation as chylomicrons. To date, only a few studies have reported the relationship between melatonin and fat absorption, only with regards to intracellular esterification [23]. Recent studies have highlighted the crucial role of NPC1L1 protein in the regulation of intestinal cholesterol absorption, and response toward different cholesterol concentrations [18]. Moreover, inhibition of NPC1L1 is known to prevent HFD-induced hepatic steatosis, weight gain, and induce a significant reduction in LDL cholesterol level, which is in alignment with the effect of melatonin under HFD [19]. Despite this significance of NPC1L1, the relationship between melatonin and various intestinal nutrient transporters including NPC1L1 has never been studied before. Our data for the first time highlights the inhibitory effect of melatonin on the expression of NPC1L1 protein under HFD (Fig. 3D). One considerable point is the expression of NPC1L1 was markedly downregulated in HFD+M. Because NPC1L1 is a critical mediator of cholesterol absorption, excessive suppression of NPC1L1 might have adverse effects caused from lack of cholesterol. However, the total amount of cholesterol transport via NPC1L1 was higher compared to extremely low expression of NPC 1 level as we can confirm in Fig. 2. It is speculated due to the extremely high content of fat amount (60%) of HFD compared to the chow diet which contains 5.54% of fat.
- The results from our assessment of various intestinal nutrient transporters collectively indicate the fact that melatonin has an inhibitory effect on intestinal nutrient absorption under circumstances where excessive calories are supplied. Thus, highlighting the regulatory role of melatonin in intestinal caloric absorption.
- The inhibitory effect of melatonin in hepatic cholesterol biosynthesis in liver has been demonstrated through previous studies [35]. However, the mechanism underlying this effect regarding the involvement of HMGCR is controversial [22]. Our examination of hepatic mRNA expression levels revealed that melatonin significantly inhibited the HFD-induced elevation of SREBP-2 and HMGCR, which promotes cholesterol biosynthesis (Fig. 4A, B). In a similar pattern, decreased expression of LDLR was noticed although not at a statistically significant level (Fig. 4C). We assume that this decrease in expression of LDLR is secondary to the decreased amount of cholesterol biosynthesis.
- Melatonin is known to exert some of its effects mediated through modulation of inflammatory cytokines or cellular interleukin responses rather than direct action on target cells [36]. Our data showed no significant differences in local and inflammatory cytokine levels between HFD and HFD+M mice (Fig. 5), which suggests the aforementioned effects of melatonin are the result of its direct action on target cells rather than indirectly mediated through circulatory factors. Additionally, we performed an in vitro cell experiment through a previously established cellular hepatic model of human hepatocytes in steatosis by treating HepG2 cells with palmitate [37,38]. Our data demonstrated decreased expression levels of SREBP-2, HMGCR, and LDLR (Fig. 6) which is consistent with our in vivo findings in mouse hepatocytes. Despite the controversial result in another in vitro study [22], our collective evidence from in vitro and in vivo mouse experiments indicates the direct action of melatonin on hepatocytes inhibiting cholesterol biosynthesis.
- This study has several limitations. First, since there are only five numbers in each group, several results were not reached statistical significance. In terms of inflammatory cytokines, the lack of significance in the HFD group might be due to the insufficient number. Second, we didn’t perform protein levels. The expression of mRNA could represent the protein expression, sometimes it may not coincide with protein expression. Although most papers related to intestinal nutrient transporters showed mRNA expression only, future studies are needed to determine how melatonin affects protein levels and activities.
- The significance of our study is uncovering the anti-obesogenic and steatoprotective mechanism of melatonin under HFD with regard to cholesterol absorption and biosynthesis. Especially, our study is the first to discover the inhibitory effect of melatonin in intestinal lipid absorption through suppression of NPC1L1, which is known to have anti-obesogenic and steatoprotective properties [19]. Moreover, we did not expose our mice to sleep deprivation to maintain the normal diurnal profile of melatonin. Our results were obtained under this circumstance which implies the therapeutic potential of melatonin as a treatment for obesity and fatty liver. Indeed, it must be noted that blood levels of melatonin were not measured in our study. Additional studies with measurements of higher serum melatonin levels in HFD+M mice and its diurnal fluctuations would further strengthen these results.
- In conclusion, our data demonstrate the inhibitory effect of melatonin in weight gain and progression of fatty liver in mice exposed to 17 weeks of HFD through suppression of lipid absorption via decreased intestinal NPC1L1 expression and decreased cholesterol synthesis.
DISCUSSION
-
CONFLICTS OF INTEREST
No potential conflict of interest relevant to this article was reported.
-
AUTHOR CONTRIBUTIONS
Conception or design: B.K. Acquisition, analysis, or interpretation of data: H.K., Y.K., B.K. Drafting the work or revising: H.K., G.L., B.K. Final approval of the manuscript: H.K., Y.K., A.L.K., G.L., Y.C., B.K.
Article information
-
Acknowledgements
- This study is funded by the beginning independent researcher program of the National Research Foundation of Korea, Project No. 2017R1C1B5017903.
- 1. World Health Organization. Obesity and overweight [Internet]. Geneva: WHO; 2021 [cited 2023 Jul 31]. Available from: https://www.who.int/news-room/fact-sheets/detail/obesity-and-overweight.
- 2. NCD Risk Factor Collaboration (NCD-RisC). Worldwide trends in body-mass index, underweight, overweight, and obesity from 1975 to 2016: a pooled analysis of 2416 population-based measurement studies in 128.9 million children, adolescents, and adults. Lancet 2017;390:2627–42.PubMedPMC
- 3. Ha KH, Kim DJ. Epidemiology of childhood obesity in Korea. Endocrinol Metab (Seoul) 2016;31:510–8.ArticlePubMedPMCPDF
- 4. Rhie S, Lee S, Chae KY. Sleep patterns and school performance of Korean adolescents assessed using a Korean version of the pediatric daytime sleepiness scale. Korean J Pediatr 2011;54:29–35.ArticlePubMedPMC
- 5. Cappuccio FP, D’Elia L, Strazzullo P, Miller MA. Sleep duration and all-cause mortality: a systematic review and meta-analysis of prospective studies. Sleep 2010;33:585–92.ArticlePubMedPMC
- 6. Kim BK, Kim BS, An SY, Lee MS, Choi YJ, Han SJ, et al. Sleep duration and glycemic control in patients with diabetes mellitus: Korea National Health and Nutrition Examination Survey 2007-2010. J Korean Med Sci 2013;28:1334–9.ArticlePubMedPMCPDF
- 7. Knutson KL, Van Cauter E. Associations between sleep loss and increased risk of obesity and diabetes. Ann N Y Acad Sci 2008;1129:287–304.ArticlePubMedPMC
- 8. Kohansieh M, Makaryus AN. Sleep deficiency and deprivation leading to cardiovascular disease. Int J Hypertens 2015;2015:615681.ArticlePubMedPMCPDF
- 9. Sato K, Meng F, Francis H, Wu N, Chen L, Kennedy L, et al. Melatonin and circadian rhythms in liver diseases: functional roles and potential therapies. J Pineal Res 2020;68:e12639.ArticlePubMedPMCPDF
- 10. Yaggi HK, Araujo AB, McKinlay JB. Sleep duration as a risk factor for the development of type 2 diabetes. Diabetes Care 2006;29:657–61.ArticlePubMedPDF
- 11. Yeo Y, Ma SH, Park SK, Chang SH, Shin HR, Kang D, et al. A prospective cohort study on the relationship of sleep duration with all-cause and disease-specific mortality in the Korean Multi-center Cancer Cohort study. J Prev Med Public Health 2013;46:271–81.ArticlePubMedPMC
- 12. Rodriguez V, Mellado C, Alvarez E, De Diego JG, Blazquez E. Effect of pinealectomy on liver insulin and glucagon receptor concentrations in the rat. J Pineal Res 1989;6:77–88.ArticlePubMed
- 13. Cipolla-Neto J, Amaral FG, Afeche SC, Tan DX, Reiter RJ. Melatonin, energy metabolism, and obesity: a review. J Pineal Res 2014;56:371–81.ArticlePubMedPDF
- 14. Obayashi K, Saeki K, Iwamoto J, Okamoto N, Tomioka K, Nezu S, et al. Exposure to light at night, nocturnal urinary melatonin excretion, and obesity/dyslipidemia in the elderly: a cross-sectional analysis of the HEIJO-KYO study. J Clin Endocrinol Metab 2013;98:337–44.ArticlePubMed
- 15. Szewczyk-Golec K, Wozniak A, Reiter RJ. Inter-relationships of the chronobiotic, melatonin, with leptin and adiponectin: implications for obesity. J Pineal Res 2015;59:277–91.ArticlePubMedPDF
- 16. She M, Laudon M, Yin W. Melatonin receptors in diabetes: a potential new therapeutical target? Eur J Pharmacol 2014;744:220–3.ArticlePubMed
- 17. Xu P, Wang J, Hong F, Wang S, Jin X, Xue T, et al. Melatonin prevents obesity through modulation of gut microbiota in mice. J Pineal Res 2017;62:e12399.ArticlePDF
- 18. Duan Y, Gong K, Xu S, Zhang F, Meng X, Han J. Regulation of cholesterol homeostasis in health and diseases: from mechanisms to targeted therapeutics. Signal Transduct Target Ther 2022;7:265.ArticlePubMedPMCPDF
- 19. Jia L, Betters JL, Yu L. Niemann-pick C1-like 1 (NPC1L1) protein in intestinal and hepatic cholesterol transport. Annu Rev Physiol 2011;73:239–59.ArticlePubMedPMC
- 20. Luo J, Yang H, Song BL. Mechanisms and regulation of cholesterol homeostasis. Nat Rev Mol Cell Biol 2020;21:225–45.ArticlePubMedPDF
- 21. Wang L, McFadden JW, Yang G, Zhu H, Lian H, Fu T, et al. Effect of melatonin on visceral fat deposition, lipid metabolism and hepatic lipo-metabolic gene expression in male rats. J Anim Physiol Anim Nutr (Berl) 2021;105:787–96.ArticlePubMedPDF
- 22. Mi Y, Tan D, He Y, Zhou X, Zhou Q, Ji S. Melatonin modulates lipid metabolism in HepG2 cells cultured in high concentrations of oleic acid: AMPK pathway activation may play an important role. Cell Biochem Biophys 2018;76:463–70.ArticlePubMedPDF
- 23. Hussain SA. Effect of melatonin on cholesterol absorption in rats. J Pineal Res 2007;42:267–71.ArticlePubMed
- 24. Michurina SV, Kolesnikov SI, Ishchenko IY, Arkhipov SA. Light-induced functional pinealectomy: expression of MT2 receptors in liver cells of C57BL/6 mice after melatonin treatment. Bull Exp Biol Med 2022;173:569–74.ArticlePubMedPDF
- 25. Chen CQ, Fichna J, Bashashati M, Li YY, Storr M. Distribution, function and physiological role of melatonin in the lower gut. World J Gastroenterol 2011;17:3888–98.ArticlePubMedPMC
- 26. Tung YT, Chiang PC, Chen YL, Chien YW. Effects of melatonin on lipid metabolism and circulating irisin in Sprague-Dawley rats with diet-induced obesity. Molecules 2020;25:3329.ArticlePubMedPMC
- 27. Keskin I, Kaplan S, Kalkan S, Sutcu M, Ulkay MB, Esener OB. Evaluation of neuroprotection by melatonin against adverse effects of prenatal exposure to a nonsteroidal anti-inflammatory drug during peripheral nerve development. Int J Dev Neurosci 2015;41:1–7.ArticlePubMedPDF
- 28. Onger ME, Kaplan S, Deniz OG, Altun G, Altunkaynak BZ, Balci K, et al. Possible promoting effects of melatonin, leptin and alcar on regeneration of the sciatic nerve. J Chem Neuroanat 2017;81:34–41.ArticlePubMed
- 29. Mirza MS. Obesity, visceral fat, and NAFLD: querying the role of adipokines in the progression of nonalcoholic fatty liver disease. ISRN Gastroenterol 2011;2011:592404.ArticlePubMedPMCPDF
- 30. Sun H, Huang FF, Qu S. Melatonin: a potential intervention for hepatic steatosis. Lipids Health Dis 2015;14:75.ArticlePubMedPMCPDF
- 31. Prunet-Marcassus B, Desbazeille M, Bros A, Louche K, Delagrange P, Renard P, et al. Melatonin reduces body weight gain in Sprague Dawley rats with diet-induced obesity. Endocrinology 2003;144:5347–52.ArticlePubMed
- 32. Shieh JM, Wu HT, Cheng KC, Cheng JT. Melatonin ameliorates high fat diet-induced diabetes and stimulates glycogen synthesis via a PKCzeta-Akt-GSK3beta pathway in hepatic cells. J Pineal Res 2009;47:339–44.PubMed
- 33. Behbodikhah J, Ahmed S, Elyasi A, Kasselman LJ, De Leon J, Glass AD, et al. Apolipoprotein B and cardiovascular disease: biomarker and potential therapeutic target. Metabolites 2021;11:690.ArticlePubMedPMC
- 34. Iqbal J, Hussain MM. Intestinal lipid absorption. Am J Physiol Endocrinol Metab 2009;296:E1183–94.ArticlePubMedPMC
- 35. Mohammadi-Sartang M, Ghorbani M, Mazloom Z. Effects of melatonin supplementation on blood lipid concentrations: a systematic review and meta-analysis of randomized controlled trials. Clin Nutr 2018;37(6 Pt A):1943–54.ArticlePubMed
- 36. Mathes AM. Hepatoprotective actions of melatonin: possible mediation by melatonin receptors. World J Gastroenterol 2010;16:6087–97.ArticlePubMedPMC
- 37. Gomez-Lechon MJ, Donato MT, Martinez-Romero A, Jimenez N, Castell JV, O’Connor JE. A human hepatocellular in vitro model to investigate steatosis. Chem Biol Interact 2007;165:106–16.ArticlePubMed
- 38. Park JY, Kim Y, Im JA, Lee H. Oligonol suppresses lipid accumulation and improves insulin resistance in a palmitateinduced in HepG2 hepatocytes as a cellular steatosis model. BMC Complement Altern Med 2015;15:185.ArticlePubMedPMCPDF
References
Figure & Data
References
Citations
- Influence of dark deprivation on the ultrastructure and mitochondrial apparatus of rat hepatocytes
D Areshidze
Morphology.2024;[Epub] CrossRef
 PubReader
PubReader ePub Link
ePub Link Cite
Cite