Inhibition of Sodium-Glucose Cotransporter-2 during Serum Deprivation Increases Hepatic Gluconeogenesis via the AMPK/AKT/FOXO Signaling Pathway
Article information

Abstract
Background
Sodium-dependent glucose cotransporter 2 (SGLT2) mediates glucose reabsorption in the renal proximal tubules, and SGLT2 inhibitors are used as therapeutic agents for treating type 2 diabetes mellitus. This study aimed to elucidate the effects and mechanisms of SGLT2 inhibition on hepatic glucose metabolism in both serum deprivation and serum supplementation states.
Methods
Huh7 cells were treated with the SGLT2 inhibitors empagliflozin and dapagliflozin to examine the effect of SGLT2 on hepatic glucose uptake. To examine the modulation of glucose metabolism by SGLT2 inhibition under serum deprivation and serum supplementation conditions, HepG2 cells were transfected with SGLT2 small interfering RNA (siRNA), cultured in serum-free Dulbecco’s modified Eagle’s medium for 16 hours, and then cultured in media supplemented with or without 10% fetal bovine serum for 8 hours.
Results
SGLT2 inhibitors dose-dependently decreased hepatic glucose uptake. Serum deprivation increased the expression levels of the gluconeogenesis genes peroxisome proliferator-activated receptor gamma co-activator 1 alpha (PGC-1α), glucose 6-phosphatase (G6pase), and phosphoenolpyruvate carboxykinase (PEPCK), and their expression levels during serum deprivation were further increased in cells transfected with SGLT2 siRNA. SGLT2 inhibition by siRNA during serum deprivation induces nuclear localization of the transcription factor forkhead box class O 1 (FOXO1), decreases nuclear phosphorylated-AKT (p-AKT), and p-FOXO1 protein expression, and increases phosphorylated-adenosine monophosphate-activated protein kinase (p-AMPK) protein expression. However, treatment with the AMPK inhibitor, compound C, reversed the reduction in the protein expression levels of nuclear p-AKT and p-FOXO1 and decreased the protein expression levels of p-AMPK and PEPCK in cells transfected with SGLT2 siRNA during serum deprivation.
Conclusion
These data show that SGLT2 mediates glucose uptake in hepatocytes and that SGLT2 inhibition during serum deprivation increases gluconeogenesis via the AMPK/AKT/FOXO1 signaling pathway.
INTRODUCTION
Two sodium-dependent glucose transporter (SGLT) family members, SGLT1 and sodium-dependent glucose cotransporter 2 (SGLT2), mediate glucose reabsorption and maintain glucose homeostasis. SGLT1 is expressed in the intestine, whereas SGLT2 is mainly expressed in the S1 and S2 segments of the renal proximal tubules [1,2]. SGLT2 inhibitors are used as antidiabetic agents to decrease plasma glucose levels by reducing the reabsorption of filtered glucose in the renal proximal tubule and increasing urinary glucose excretion [3,4]. Recent studies reported the expression of SGLT2 in hepatocytes and in the liver [5,6]. However, the role of SGLT2 in the hepatocytes remains unclear.
Safety issues associated with newer glucose-lowering medications used in patients with type 2 diabetes mellitus (T2DM) are important. Hypoglycemia is when the body’s blood glucose or sugar levels are lower than normal and can lead to coma or even death [7,8]. Antidiabetic medications such as meglitinides and sulfonylureas cause hypoglycemia by stimulating the release of insulin from pancreatic beta cells [9], whereas SGLT2 inhibitors are a better option for improving glycemic control with a low risk of hypoglycemia, even when co-administered with insulin [10].
A trial using SGLT2 inhibitors in patients with T2DM reported that dapagliflozin improved insulin sensitivity by lowering plasma glucose and increased gluconeogenesis by increasing the glucagon/insulin ratio [11]. Gluconeogenesis is a metabolic process synthesizing glucose from non-carbohydrate precursors when insufficient or absent dietary intake [12]. During the first 8 hours of fasting, glucose is provided through glycogenolysis, the process of glycogen breakdown in the liver and muscles; when stored glycogen is depleted, glucose is synthesized to maintain homeostasis by increasing gluconeogenesis [13,14]. Gluconeogenesis occurs predominantly in the liver and kidney; during starvation, the liver accounts for 80% to 90% of gluconeogenesis, and the kidney accounts for 10% to 20% [15,16]. Furthermore, gluconeogenesis in the liver is regulated by the activation of key enzymes such as glucose 6-phosphatase (G6pase), fructose 1,6-bisphosphatase (Fbpase), pyruvate carboxylase, and phosphoenolpyruvate carboxykinase (PEPCK).
Forkhead box class O (FOXO) transcription factor family members, including FOXO1, FOXO3a, FOXO4, and FOXO6, play important roles in regulating cellular processes, including cell proliferation, stress resistance, and metabolism [17]. FOXO is phosphorylated by several protein kinases, resulting in its translocation from the nucleus to the cytoplasm, modification of DNA-binding affinity, and alteration of the transcriptional activity of target genes [18,19]. In particular, FOXO1 is closely associated with hepatic gluconeogenesis, and FOXO1 expression is upregulated during fasting and energy starvation [20,21].
In the present study, we investigated the role and regulatory mechanisms of SGLT2 inhibition in maintaining glucose homeostasis in hepatocytes under serum deprivation and serum supplementation conditions.
METHODS
Cell culture
Huh7 and HepG2 human hepatoma cell lines were purchased from American Type Culture Collection (ATCC, Manassas, VA, USA). Cells were cultured in Dulbecco’s modified Eagle’s medium (DMEM) high-glucose (25 mmol/L) (Hyclone, Logan, UT, USA) supplemented with 10% fetal bovine serum (FBS; Hyclone) and 1% penicillin/streptomycin (Gibco, Thermo Fisher Scientific Inc., Waltham, MA, USA), and incubated in an incubator at 37°C with 5% CO2. All experiments in this study were performed in HepG2 cells, except that the glucose uptake assay was performed in Huh7 cells. HepG2 cells have the characteristic of forming aggregation, so Huh7 cells were used to compare clear differences in the short-term glucose uptake assay.
Glucose uptake
Glucose uptake was assessed using a glucose uptake cell-based assay kit (Cayman Chemical, Ann Arbor, MI, USA) according to the manufacturer’s instructions. Huh7 cells were cultured in sodium (Na+) buffer, including 140 mM NaCl, 5 mM KCl, 2.5 mM CaCl2, 1 mM MgSO4, 1 mM KH2PO4, and 10 mM HEPES, or in sodium-free (Na+-free) buffer, including 140 mM N-methyld-glucamine instead of NaCl, for 1 hour [22]. Cells were treated with 10 nM empagliflozin and dapagliflozin (Cayman Chemical) in glucose-free DMEM low-glucose (5 mmol/L) for 24 hours to determine the change of glucose uptake by SGLT2 inhibitor in hepatocytes. Then, the cell culture medium was replaced by sodium buffer and exposed to 200 μg/mL 2-(N-(7-nitrobenz-2-oxa1,3-diazol-4-yl)amino)-2-deoxyglucose (2-NBDG), a fluorescently labeled deoxyglucose analog, for 1 hour, washed and imaged by fluorescence microscope (IX71, Olympus, Tokyo, Japan). Moreover, cells were lysed with 0.1 M NaOH, neutralized with 0.1 M HCl, and measured using a microplate reader (excitation/emission=485/535 nm).
Cell viability assay
HepG2 cell viability was detected using Cell Counting Kit-8 (CCK-8) colorimetric assay (Abcam, Cambridge, UK), according to the manufacturer’s instructions. Briefly, cells were cultured in 96-well plate, then 10 μL of the CCK-8 solution was added to each well and incubated for 2 hours at 37°C. Absorbance was measured using a microplate reader at 450 nm (Tecan, Grödig, Austria).
Immunofluorescence staining
HepG2 cells cultured on coverslips were fixed with 4% formaldehyde for 10 minutes, permeabilized with 0.25% Triton X-100, and blocked with a normal goat serum solution for 1 hour. The cells were then incubated overnight with the primary anti-FOXO1 or anti-PEPCK antibody at 4°C and were incubated with an Alexa Fluor 488 secondary antibody or an Alexa Fluor 546 secondary antibody (Invitrogen, Thermo Fisher Scientific, Waltham, CA, USA) for 1 hour at room temperature. After washing with phosphate-buffered saline, the cells were mounted using an aqueous mounting medium with 4′,6-diamidino-2-phenylindole (DAPI) (ab104139, Abcam), and images were taken using a fluorescence microscope (BX51, Olympus).
Quantitative real-time polymerase chain reaction
Total RNA was extracted from HepG2 cells using TRIzol reagent (Invitrogen) and quantified using spectrophotometry (Nanodrop, Thermo Scientific). cDNA was synthesized using a High-Capacity RNA-to-cDNA Kit (Applied Biosystems, Foster City, CA, USA), and the expression of target genes was measured using a LightCycler 480 SYBR Green (Roche, Lewis, UK). The primer sequences used for polymerase chain reaction (PCR) are listed in Supplemental Table S1. Thermocycling conditions of quantitative real-time PCR were as follows: 45 cycles of denaturation at 95°C for 30 seconds, 55°C for 30 seconds, extension at 72°C for 30 seconds, and a final extension at 72°C for 5 minutes. β-Actin was used as the reference control to normalize the relative expression levels of the target genes.
Western blotting
Total proteins were extracted from cultured cells using radioimmunoprecipitation assay (RIPA) buffer (Cell Signaling Technology, Danvers, MA, USA) containing Halt Protease and Phosphatase Inhibitor Cocktail (100×) (Thermo Fisher Scientific, Rockford, IL, USA). Nuclear and cytoplasmic protein fractions were isolated from cultured cells using a nuclear extraction kit (Cayman Chemical). Cell lysates containing 20 µg protein were mixed with 4× lithium dodecyl sulfate sample buffer and 10× reducing sample agent (Invitrogen), boiled at 95°C for 10 minutes. Protein samples were loaded onto a 4% to 12% Bis-Tris NuPAGE gel (Thermo Fisher Scientific) and transferred to a polyvinylidene difluoride (PVDF) membrane using an iBlot2 PVDF stack (Invitrogen). The membranes were blocked with 5% bovine serum albumin in tris-buffered saline with Tween-20 for 1 hour at room temperature, following which they were incubated overnight at 4°C with the following specific primary antibodies (Supplemental Table S2). Membranes were washed and incubated with horseradish peroxidase-conjugated secondary antibodies for 1 hour at room temperature. Protein bands were visualized using an enhanced chemiluminescence reagent (DAWINBio, Hanam, Korea), and band intensities were quantified using ImageJ software (National Institutes of Health, Bethesda, MD, USA).
Transient small interfering RNA transfection
The cells were transfected with SGLT2 small interfering RNA (siRNA) to determine the modulation of glucose metabolism by SGLT2 inhibition under serum deprivation and serum supplementation conditions. siRNA duplexes against SGLT2 and the negative control were purchased from Bioneer (Daejeon, Korea), and the cells were transfected with SGLT2 siRNA using Lipofectamine RNAiMAX transfection reagent (Invitrogen), according to the manufacturer’s instructions. Cells pre-transfected with SGLT2 siRNA were cultured in serum-free DMEM (25 mmol/L glucose) for 16 hours and then in media supplemented with or without 10% FBS for 8 hours.
Statistical analysis
All experiments were performed in triplicate, and data are presented as mean±standard error values. Statistical comparisons between groups were conducted using the SPSS software version 12.0 (SPSS Inc., Chicago, IL, USA). The data were analyzed using a one-way analysis of variance, followed by Bonferroni post hoc pairwise comparisons to calculate significant differences between the mean values obtained from the different experimental groups. P<0.05 was considered statistically significant.
RESULTS
SGLT2 inhibitors reduce glucose uptake in hepatocytes
SGLT2, mainly present in the renal proximal tubule, maintains glucose homeostasis by regulating glucose reabsorption in a sodium-dependent manner [3]. Huh7 cells were treated with various concentrations of the SGLT2 inhibitors, empagliflozin and dapagliflozin, to determine the direct role of SGLT2 in regulating hepatic glucose metabolism. Empagliflozin and dapagliflozin decreased the glucose uptake dose-dependently (Fig. 1). Glucose uptake increased in cells treated with sodium buffer compared to those treated with sodium-free buffer. Interestingly, however, the expression of glucose transporter 2 (GLUT2), another glucose transporter in the liver, was increased in the presence of glucose but was not regulated by SGLT2 inhibitors (Supplemental Fig. S1). These results indicate that SGLT2 plays a role in hepatic glucose metabolism and that SGLT2 inhibitors may play a role in directly inhibiting glucose uptake in hepatocytes without affecting GLUT2.
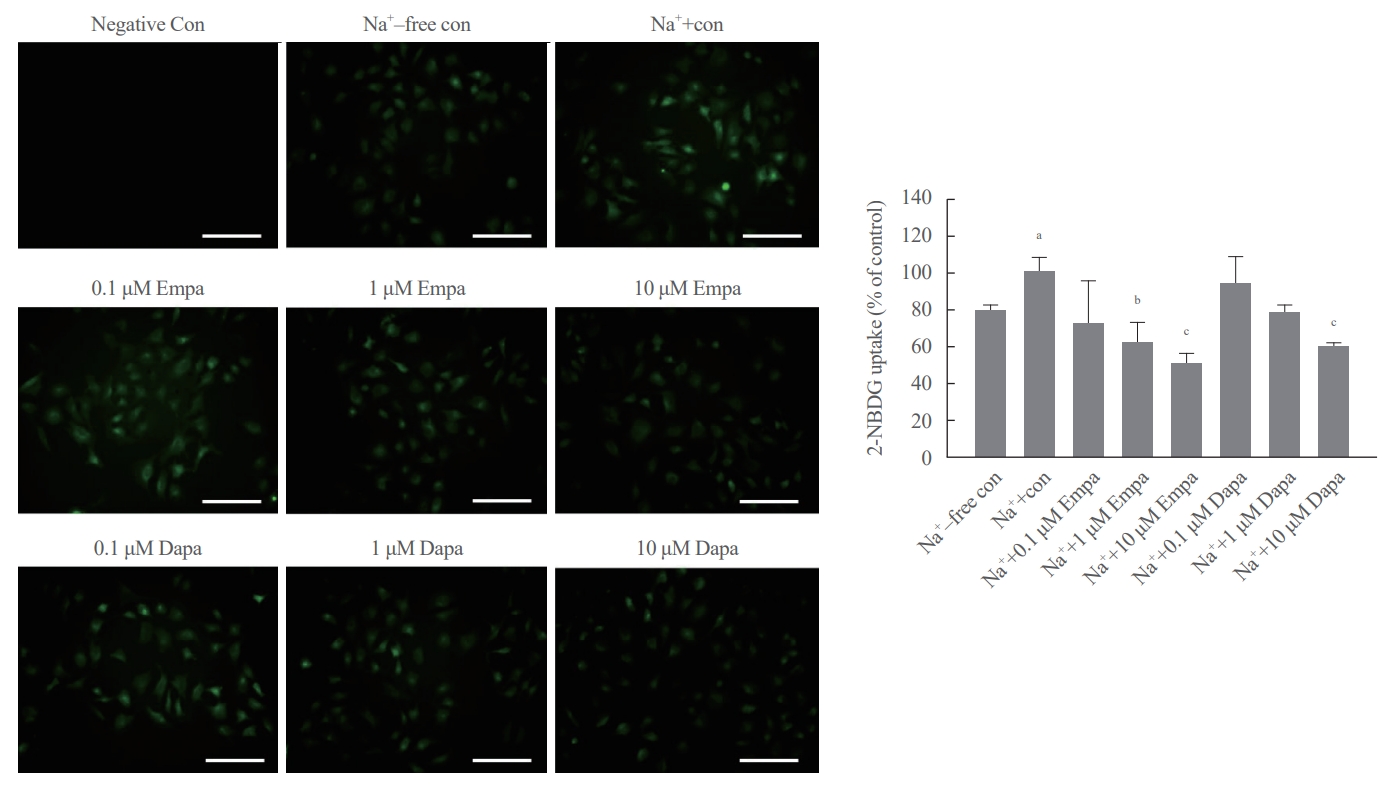
Empagliflozin (Empa) and dapagliflozin (Dapa) inhibit hepatic glucose uptake in a sodium (Na+) dependent manner. Huh7 cells are cultured in sodium-free (Na+-free) buffer or sodium (Na+) buffer and treated with various concentrations of the SGLT2 inhibitors Empa and Dapa. Glucose uptake is determined using a glucose uptake assay kit. The cells were photographed using a fluorescence microscope (magnification, ×400); scale bars=20 μm. Con, control; 2-NBDG, 2-(N-(7-nitrobenz-2-oxa-1,3-diazol-4-yl)amino)-2-deoxyglucose. aP<0.05 when compared with sodium-free buffer; bP<0.05 and cP<0.01 when compared with sodium-supplemented buffer.
SGLT2 inhibition increases gluconeogenesis in HepG2 during serum deprivation
Glucose deprivation induces cell death. Cells pre-transfected with SGLT2 siRNA were cultured under serum deprivation (serum-free media for 24 hours) and serum supplementation (serum-free media for 16 hours+serum-supplemented media for 8 hours) conditions to determine the effects of SGLT2 inhibition on serum starved cells. SGLT2 inhibition by siRNA in HepG2 cells did not lead to a compensatory increase in GLUT2 gene expression, but increased the expression of sirtuin 1 (SIRT1), whose expression is increased during starvation, and gluconeogenesis-associated genes, including peroxisome proliferator-activated receptor gamma co-activator 1 alpha (PGC-1α), PEPCK, and G6pase under serum deprivation conditions (Fig. 2). In addition, SGLT2 inhibition during serum deprivation decreased the expression of extracellular signal-regulated kinase (ERK), the c-Jun N-terminal kinases (JNK), and proliferating cell nuclear antigen (PCNA), which are involved in cell proliferation (Supplemental Fig. S2A). Nevertheless, the expression of cleaved caspase-3 protein did not change between the groups, and serum deprivation for 24 hours in cells pre-transfected with SGLT2 siRNA did not induce cell death (Supplemental Fig. S2A, B). These results indicate that SGLT2 inhibition under serum deprivation conditions decreases cell proliferation and increases gluconeogenesis for cell survival.
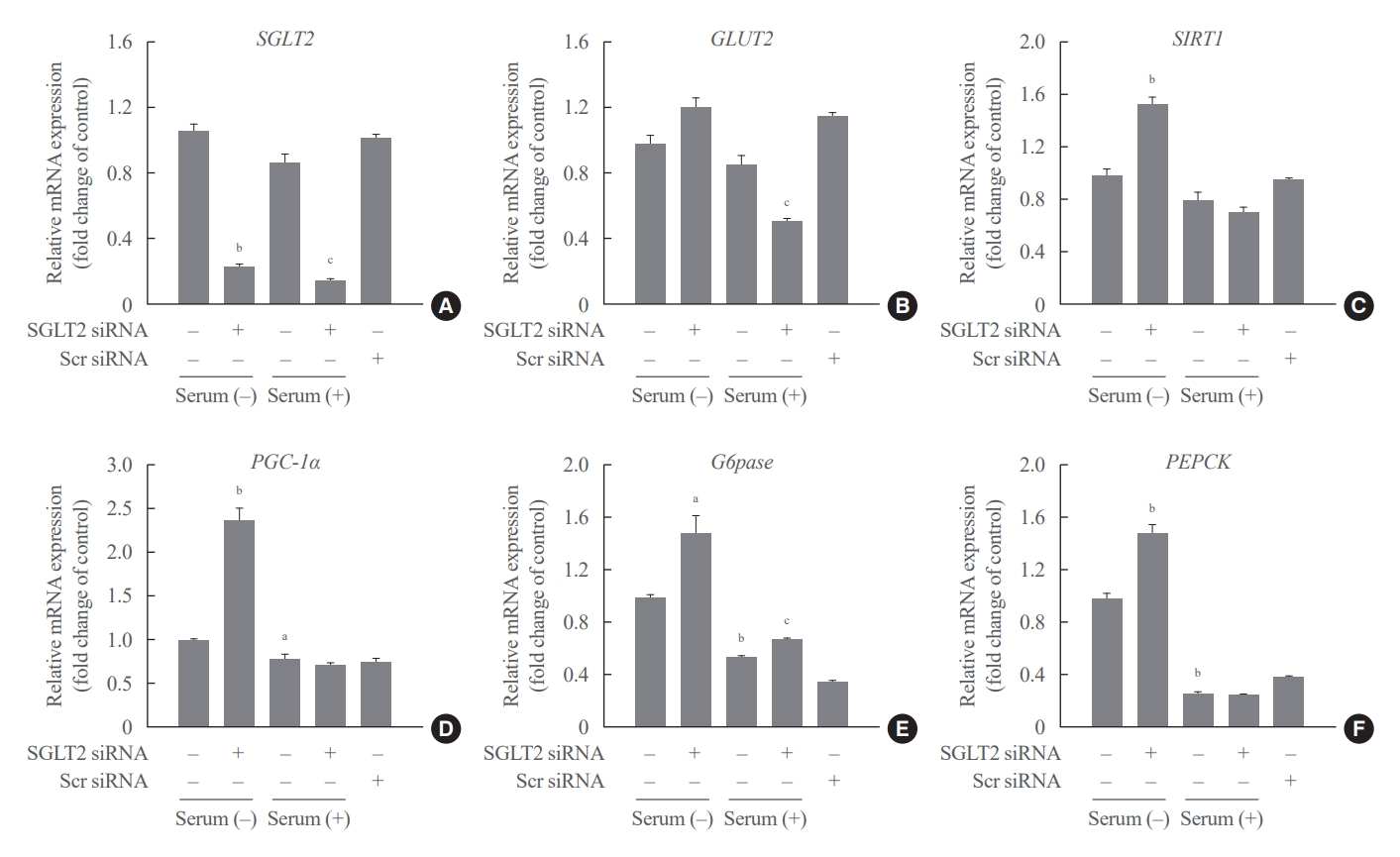
Sodium-dependent glucose cotransporter 2 (SGLT2) inhibition during serum deprivation increases the gluconeogenesis-associated genes’ expression in HepG2 cells. HepG2 cells pre-transfected with 50 nM SGLT2 small interfering RNA (siRNA) are cultured in serumfree Dulbecco’s modified Eagle’s medium (DMEM) (25 mmol/L glucose) for 16 hours and then in media supplemented with (serum supplementation) or without (serum deprivation) 10% fetal bovine serum for 8 hours. The mRNA expression levels of the genes encoding (A) SGLT2, (B) glucose transporter 2 (GLUT2), (C) sirtuin 1 (SIRT1), (D) peroxisome proliferator-activated receptor gamma co-activator 1 alpha (PGC-1α), (E) glucose 6-phosphatase (G6pase), and (F) phosphoenolpyruvate carboxykinase (PEPCK) are analyzed with quantitative real-time polymerase chain reaction and normalized to the β-Actin gene. Scr, scrambled. aP<0.05 and bP<0.01 when compared with the serum-free control cells; cP<0.01 when compared with the serum-supplemented control cells.
SGLT2 inhibition increases the expression of p-AMPK and decreases the expression of nuclear p-AKT and p-FOXO1
We examined whether SGLT2 inhibition during serum deprivation and serum supplementation regulates the expression of FOXO1, a transcription factor involved in hepatic gluconeogenesis. AKT in the nucleus phosphorylates FOXO1, translocates FOXO1 to the cytoplasm, and inhibits its transcription activities [23,24]. SGLT2 inhibition by siRNA during serum deprivation, but not during serum supplementation, increased the expression of phosphorylated-adenosine monophosphate-activated protein kinase (p-AMPK) and decreased the expression of nuclear p-AKT and p-FOXO1 (Fig. 3A, B). In addition, SGLT2 inhibition by siRNA increased the nuclear localization of FOXO1, demonstrated by immunofluorescence staining (Fig. 3C). These results in cells transfected with SGLT2 siRNA were also consistently seen in cells treated with SGLT2 inhibitors, empagliflozin and dapagliflozin (Supplemental Fig. S3).
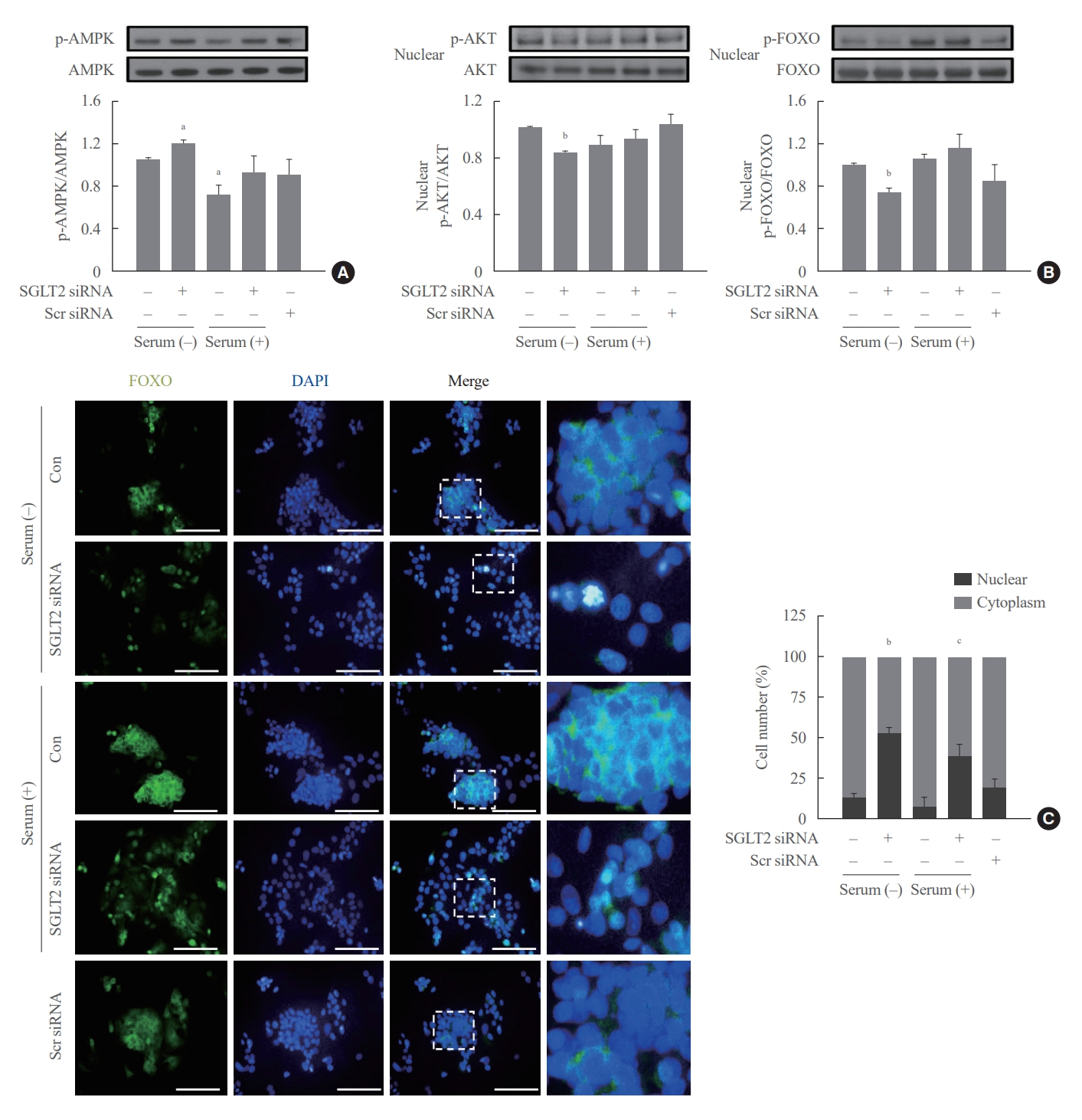
Sodium-dependent glucose cotransporter 2 (SGLT2) inhibition during serum deprivation increases the expression of phosphorylatedadenosine monophosphate-activated protein kinase (p-AMPK) and decreases the expression of nuclear p-AKT and phosphorylated-forkhead box class O 1 (p-FOXO1) proteins in HepG2 cells. HepG2 cells pre-transfected with 50 nM SGLT2 small interfering RNA (siRNA) are cultured in serum-free Dulbecco’s modified Eagle’s medium (25 mmol/L glucose) for 16 hours and then cultured in media supplemented with (serum supplementation) or without (serum deprivation) 10% fetal bovine serum for 8 hours. (A, B) Expression levels of p-AMPK, p-AKT, and p-FOXO1 are analyzed using Western blotting. (C) Subcellular localization of FOXO1 is determined by performing immunofluorescence staining, and the cells were photographed using a fluorescence microscope (magnification, ×400); scale bars=100 μm. Scr, scrambled; DAPI, 4′,6-diamidino-2-phenylindole; Con, control. aP<0.05 and bP<0.01 when compared with the serum-free control cells; cP<0.01 when compared with the serum-supplemented control cells.
AMPK inhibition in HepG2 cells transfected with SGLT2 siRNA under serum deprivation increases the expression of nuclear p-AKT and p-FOXO1 and reduces the expression of PEPCK
To determine whether AMPK increased by SGLT2 inhibition modulates AKT-FOXO1 signaling, cells pre-transfected with SGTL2 siRNA were treated with the AMPK inhibitor compound C. Inhibition of AMPK by compound C treatment decreased the expression of p-AMPK and PEPCK proteins and increased the expression of nuclear p-AKT and p-FOXO1 proteins (Fig. 4A, B). In addition, the increase in p-AMPK and PEPCK protein expression and decrease in nuclear p-AKT and p-FOXO1 protein expression in cells transfected with SGLT2 siRNA were reversed by treatment with compound C. Consistently, immunofluorescence staining for PEPCK demonstrated that the increased expression of PEPCK in cells transfected with SGLT2 siRNA was reduced by compound C (Fig. 4C). These data show that AMPK, increased by SGLT2 inhibition during serum deprivation, increases hepatic gluconeogenesis through the AKT-FOXO1 pathway.
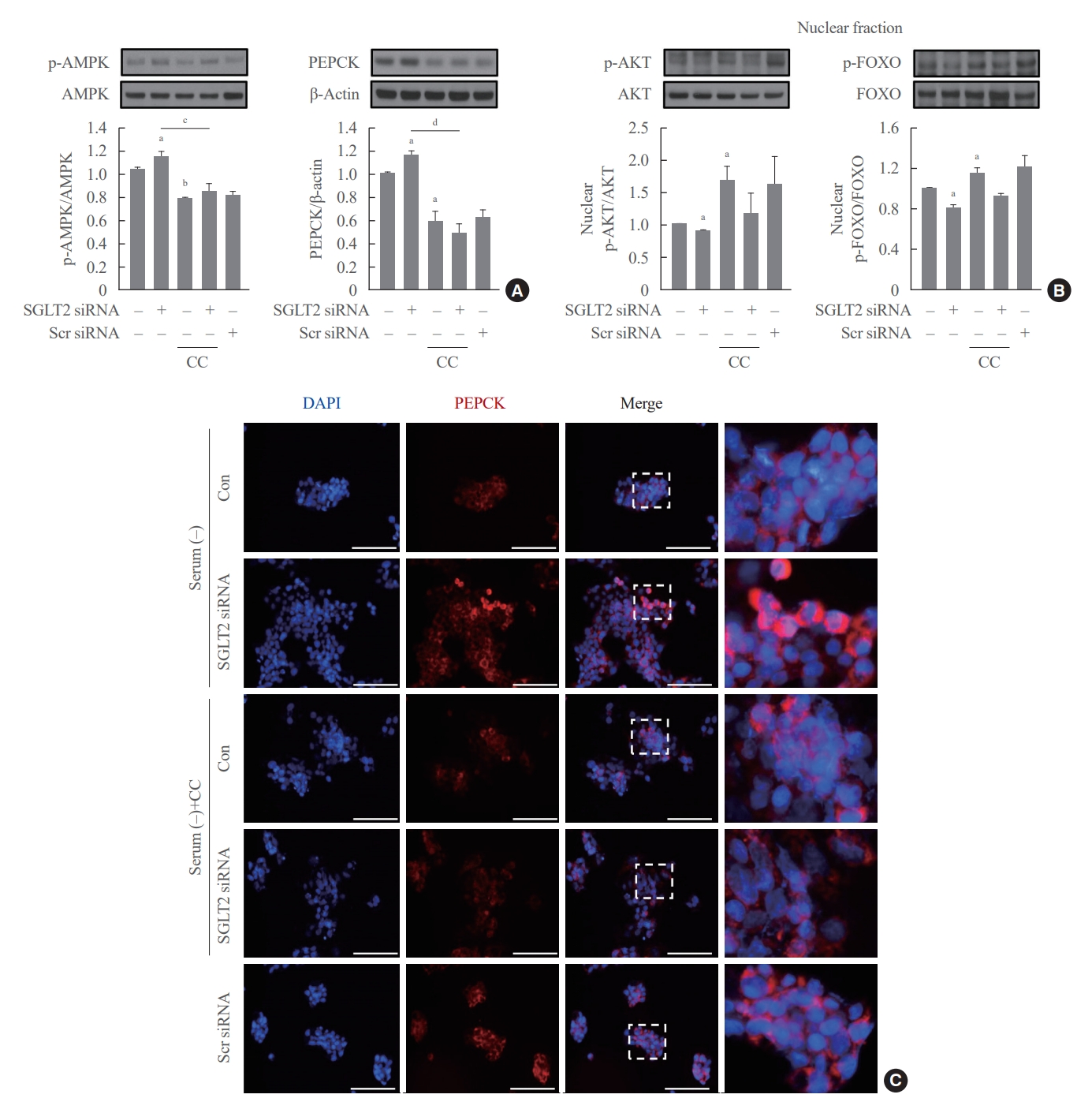
Adenosine monophosphate-activated protein kinase (AMPK) inhibition in cells transfected sodium-dependent glucose cotransporter 2 (SGLT2) small interfering RNA (siRNA) during serum deprivation decreases the expression of p-AMPK and phosphoenolpyruvate carboxykinase (PEPCK) proteins and increases the expression of nuclear p-AKT and phosphorylated-forkhead box class O 1 (p-FOXO1) proteins. HepG2 cells pre-transfected with 50 nM SGLT2 siRNA in serum-free Dulbecco’s modified Eagle’s medium are treated with 10 µM of AMPK inhibitor compound C (CC). (A, B) Expression levels of p-AMPK, PEPCK, p-AKT, and p-FOXO1 are analyzed using Western blotting. (C) The presence of PEPCK is determined by performing immunofluorescence staining, and the cells were photographed using a fluorescence microscope (magnification, ×400); scale bars=100 μm. Scr, scrambled; DAPI, 4′,6-diamidino-2-phenylindole; Con, control. aP<0.05 and bP<0.01 when compared with the serum-free control cells; cP<0.05 and dP<0.01 when compared with the cells transfected with SGLT2 siRNA without CC.
DISCUSSION
In this study, we determined that SGLT2 inhibition during serum deprivation, but not during serum supplementation, increased hepatic gluconeogenesis via the AMPK/AKT/FOXO1 signaling pathway to protect against hypoglycemia-induced apoptosis (Fig. 5). The SGLT2 inhibitors, empagliflozin and dapagliflozin, decreased the sodium-dependent glucose uptake in Huh7 cells. SGLT2 inhibition under serum deprivation conditions increased the expression levels of gluconeogenesis-associated genes and AMPK protein and reduced the expression of nuclear p-AKT and p-FOXO1 proteins. In addition, nuclear localization of FOXO1, a major transcription factor for hepatic gluconeogenesis, was observed in cells transfected with SGLT2 siRNA during serum deprivation. However, in cells transfected with SGLT2 siRNA during serum deprivation, treatment with the AMPK inhibitor compound C reversed the decrease in the expression of nuclear p-AKT and p-FOXO1 and decreased the expression of PEPCK.
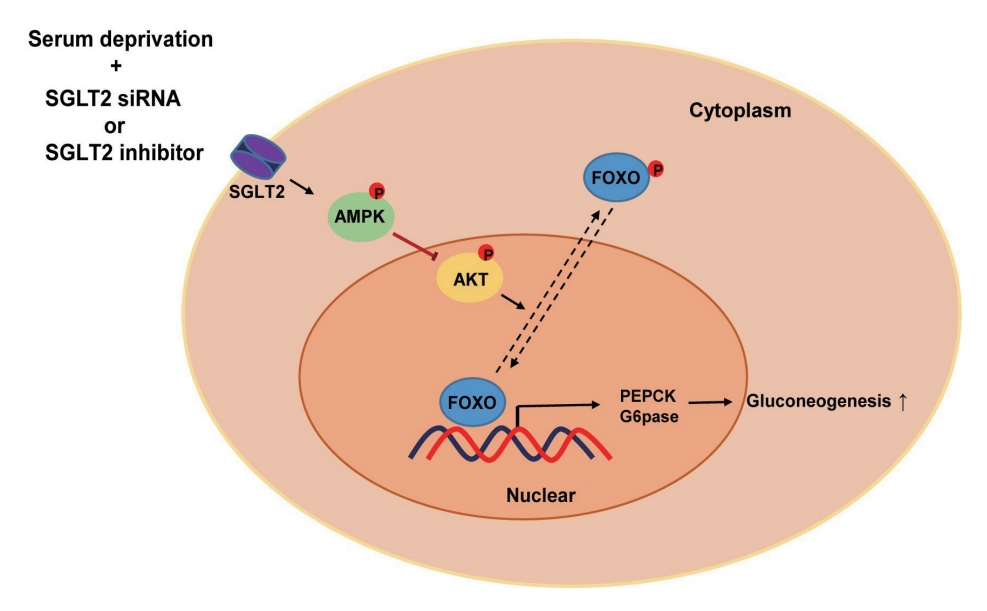
Schematic diagram illustrating a possible mechanism: sodium-dependent glucose cotransporter 2 (SGLT2) inhibition during serum deprivation stimulates hepatic gluconeogenesis. Increased adenosine monophosphate-activated protein kinase (AMPK) by SGLT2 inhibition reduced the expression of nuclear p-AKT and phosphorylated-forkhead box class O 1 (p-FOXO1), consequently increasing gluconeogenesis-related gene expression. siRNA, small interfering RNA; PEPCK, phosphoenolpyruvate carboxykinase; G6pase, glucose 6-phosphatase.
The liver plays a vital role in the regulation of glucose homeostasis. Excessive glucose in the liver is used as a substrate for fatty acid synthesis, causing fatty liver disease, whereas glucose deprivation in hepatocytes induces growth arrest and cell death [1,25]. Gluconeogenesis is stimulated by glucagon, growth hormone, epinephrine, and cortisol, which regulate transcription factors for gluconeogenesis such as cAMP response element binding protein (CREB) and FOXO and transcriptional co-activator such as CREB binding protein (CBP)/p300, CREB regulated transcription co-activator 2 (CRTC2), and PGC-1α [26,27]. During hypoglycemia, glucagon, a hormone secreted by pancreatic α cells, releases glucose from the liver into the blood by activating glycogenolysis and gluconeogenesis [28]. In patients with T2DM, the SGLT2 inhibitor dapagliflozin improves insulin sensitivity by reducing plasma glucose concentrations via glycosuria and increases endogenous glucose production by increasing the glucagon/insulin ratio [11]. Bonner et al. [29] reported that dapagliflozin promotes glucagon secretion from pancreatic alpha cells and hepatic gluconeogenesis. In addition, even when combined with insulin, SGLT2 inhibitors improve glycosylated hemoglobin A1c levels, fasting plasma glucose levels, and body weight without increasing the risk of hypoglycemia [30]. In this study, we observed that despite the absence of glucagon, SGLT2 inhibition in hepatocytes increased the expression of gluconeogenesis-associated genes during serum deprivation. These results indicate that serum starvation would aggravate energy deficiency in cells transfected with SGLT2 siRNA, which blocks glucose uptake, and that SGLT2 inhibition directly prevents hypoglycemia by increasing hepatic gluconeogenesis during serum deprivation.
FOXO proteins are transcription factors that play significant roles in autophagy, cell death, stress resistance, cell viability, metabolism, and differentiation [31]. Aoyama et al. [32] reported that feeding promotes the phosphorylation and nuclear exclusion of Foxo1 and reduces the expression levels of gluconeogenesis-associated genes such as G6pase, PEPCK, and PGC-1 in C57BL/6 mice. Whereas FOXO1 translocates to the nucleus during fasting and promotes the transcription of G6pase and PEPCK genes in the liver [26,33]. Phosphorylation of FOXO by AKT induces its translocation to the cytosol, thereby suppressing the transcriptional function of FOXO [23,34]. Under glucose deprivation, AMPK plays an opposite role to that of AKT in protecting cells from metabolic stress, and the AMPK agonists 5-aminoimidazole-4-carboxamide ribonucleoside (AICAR) and phenformin dephosphorylate AKT [35]. In this study, AMPK expression was increased in cells transfected with SGLT2 siRNA during serum deprivation. AMPK inhibitor treatment reversed the decreased expression of nuclear p-AKT and p-FOXO1 and decreased the expression of cytoplasmic PEPCK in cells transfected with SGLT2 siRNA during serum deprivation. Thus, these data suggest that AMPK increased by SGLT2 inhibition during serum deprivation stimulates hepatic gluconeogenesis through the AKT-FOXO1 signaling pathway to protect cells from energy deprivation-induced stress.
The regulatory effect of AMPK on FOXO-mediated gluconeogenesis is controversial. Guo et al. [36] reported that metformin, as an AMPK agonist, decrease FOXO1 S273 by inhibiting protein kinase A (PKA), blocks FOXO nuclear translocation, suppressing gluconeogenesis, in cells treated with glucagon. However, FOXO has multiple phosphorylation sites, and the nuclear-cytoplasmic translocation of FOXO depends on the activation of each phosphorylation site. In this study, we observed that AMPK activated by SGLT2 inhibition during serum deprivation, decreased phospho-FOXO1 S256 by inhibiting AKT and increased gluconeogenesis. FOXO1 is phosphorylated at T24, S256, and S319 by AKT that is followed by nuclear exclusion [37]. Thus, the regulation of gluconeogenesis by AMPK through FOXO depends on the phosphorylation site of FOXO and the subsequent localization of FOXO. Therefore, the conflicting results in the two studies may be due to differences in the mechanism activating FOXO as a transcription factor and the experimental conditions (glucagon treatment vs. serum deprivation condition).
Two families of glucose transporters regulate glucose uptake: GLUTs and SGLTs. The GLUT family contains 14 members and is expressed in a tissue-specific manner. GLUT2 is the main glucose-facilitating transporter isoform in the liver that controls glucose uptake depending on the circulating glucose concentration [38,39]. The role of SGLT2 in mediating the active transport of glucose is sodium-dependent. In this study, glucose uptake was reduced in a sodium-free medium compared to that in a sodium medium, indicating that GLUT2 regulates sodium-independent glucose uptake. Interestingly, no compensatory increase in GLUT2 expression was observed in cells treated with SGLT2 inhibitors or in cells transfected with SGLT2 siRNA compared to control cells. In addition, the inhibition of SGLT2 during serum deprivation further reduced the expression of cell proliferation-related factors, such as EKR, JNK, and PCNA, compared to serum supplementation. The mitogen-activated protein (MAP) kinase pathway, mediated by ERK, JNK, and p38 protein kinases, regulates proliferation, differentiation, development, transformation, and apoptosis [40]. PCNA, a cell proliferation marker, is a downstream factor of ERK and PCNA expression is down-regulated by MAPK inhibitor or siRNA [41]. Thus, these results suggest that GLUT2 cannot replace SGLT2, and SGLT2 inhibition during serum deprivation, compared to serum supplementation, seems to minimize energy expenditure by inhibiting cell proliferation.
In conclusion, the present study showed that SGLT2 plays a role in regulating glucose uptake in hepatocytes and that inhibition of SGLT2 during serum deprivation protects against starvation-induced cell damage by increasing hepatic gluconeogenesis. Therefore, SGLT2 inhibition may be a safe approach for lowering blood glucose levels in patients with diabetes and simultaneously improving liver disease associated with high blood glucose levels.
Supplementary Material
Supplemental Table S1.
List of Human Primers for Quantitative Real-Time Polymerase Chain Reaction
Supplemental Table S2.
List of Primary Antibodies Used for Western Blotting
Supplemental Fig. S1.
Glucose transporter 2 (GLUT2) expression increases in the presence of glucose but is not regulated by sodium-dependent glucose cotransporter 2 inhibitors. Cells were treated with 10 nM empagliflozin (Em) and dapagliflozin (Da) in low-glucose (LG; 5.5 mmol/L) Dulbecco’s modified Eagle’s medium (DMEM) and high-glucose (HG; 25 mmol/L) DMEM for 24 hours. The expression of GLUT2 gene was analyzed using quantitative real-time polymerase chain reaction and normalized to the β-Actin gene. Con, control. aP<0.01 when compared with each groups cultured in LG.
Supplemental Fig. S2.
Sodium-dependent glucose cotransporter 2 (SGLT2) inhibition during serum deprivation decreases cell proliferation but does not induce cell death. HepG2 cells pre-transfected with 50 nM SGLT2 small interfering RNA (siRNA) are cultured in serum-free Dulbecco’s modified Eagle’s medium (25 mmol/L glucose) for 16 hours and then in media supplemented with (serum supplementation) or without (serum deprivation) 10% fetal bovine serum for 8 hours. (A) Expression levels of phosphorylated-extracellular signal-regulated kinase (p-ERK), phosphorylated-c-Jun N-terminal kinase (p-JNK), proliferating cell nuclear antigen (PCNA), and cleaved caspase 3 are analyzed using Western blotting. (B) Cell viability was mesured using Cell Counting Kit-8 assay. Scr, scrambled. aP<0.05 and bP<0.01 when compared with the serum-free control cells.
Supplemental Fig. S3.
Sodium-dependent glucose cotransporter 2 (SGLT2) inhibitor treatment during serum deprivation increases the expression of phosphorylated-adenosine monophosphate-activated protein kinase (p-AMPK) and decreases the expression of nuclear p-AKT and phosphorylated-forkhead box class O 1 (p-FOXO1) proteins in HepG2 cells. HepG2 cells pre-treated with 10 nM empagliflozin (Empa) and dapagliflozin (Dapa) are cultured in serum-free Dulbecco’s modified Eagle’s medium (25 mmol/L glucose) for 16 hours and then cultured in media supplemented with (serum supplementation) or without (serum deprivation) 10% fetal bovine serum for 8 hours. (A, B) Expression levels of p-AMPK, p-AKT, and p-FOXO1 are analyzed using Western blotting. (C) Subcellular localization of FOXO1 is determined by performing immunofluorescence staining, and the cells were photographed using a fluorescence microscope (magnification, ×400); scale bars=100 μm. Con, control; DAPI, 4′,6-diamidino-2-phenylindole.
Notes
CONFLICTS OF INTEREST
Won-Young Lee is an editor-in-chief and Eun-Jung Rhee is a deputy editor of the journal. But they were not involved in the peer reviewer selection, evaluation, or decision process of this article. No other potential conflicts of interest relevant to this article were reported.
AUTHOR CONTRIBUTIONS
Conception or design: J.L., S.W.H., S.J.M., E.J.R., W.Y.L. Acquisition, analysis, or interpretation of data: J.L., S.W.H., M.J.K., Y.M.L., S.J.M., H.K., S.E.P., E.J.R., W.Y.L. Drafting the work or revising: J.L., S.W.H., M.J.K., Y.M.L., S.J.M., H.K., S.E.P., E.J.R., W.Y.L. Final approval of the manuscript: J.L., S.W.H., E.J.R., W.Y.L.
Acknowledgements
This study was supported by the National Research Foundation (NRF) funded by the Korean government (NRF-2022R1F1A 1072882) (http://www.nrf.re.kr). The funders played no role in the study design, data collection, analysis, decision to publish, or preparation of the manuscript.