Serotonergic Regulation of Hepatic Energy Metabolism
Article information

Abstract
The liver is a vital organ that regulates systemic energy metabolism and many physiological functions. Nonalcoholic fatty liver disease (NAFLD) is the commonest cause of chronic liver disease and end-stage liver failure. NAFLD is primarily caused by metabolic disruption of lipid and glucose homeostasis. Serotonin (5-hydroxytryptamine [5-HT]) is a biogenic amine with several functions in both the central and peripheral systems. 5-HT functions as a neurotransmitter in the brain and a hormone in peripheral tissues to regulate systemic energy homeostasis. Several recent studies have proposed various roles of 5-HT in hepatic metabolism and inflammation using tissue-specific knockout mice and 5-HT-receptor agonists/antagonists. This review compiles the most recent research on the relationship between 5-HT and hepatic metabolism, and the role of 5-HT signaling as a potential therapeutic target in NAFLD.
INTRODUCTION
The liver is an essential organ that regulates systemic energy metabolism and several physiological functions, including major macronutrient metabolism, immunomodulation, lipid metabolism and cholesterol homeostasis, and detoxification of several harmful reactive chemical species [1]. Thus, hepatic dysfunction is associated with systemic diseases such as cardiovascular disease, chronic kidney disease, obesity, and type 2 diabetes mellitus [2].
Nonalcoholic fatty liver disease (NAFLD), the commonest cause of chronic liver disease and end-stage liver failure [3], is defined as the accumulation of excess fat in the liver and includes a wide range conditions from simple fat accumulation in hepatic cells to nonalcoholic steatohepatitis, liver cirrhosis, and, finally, liver cancer [2]. Thus, NAFLD is a multisystem disease that is associated with both cardiovascular and hepatic morbidity and mortality [2,4]. The development and progression of NALFD are associated with multiple pathogenic mechanisms, such as fatty acid accumulation due to high lipid intake, insulin resistance-related upregulation of lipogenesis, lipid peroxidation-mediated liver injury, and elevated oxidative or endoplasmic reticulum stress [5,6], and metabolic disruption of lipid and glucose homeostasis is a major contributor of NAFLD pathogenesis [5]. Therefore, the restoration of hepatic metabolic homeostasis is a promising therapeutic approach for the treatment of NAFLD.
Serotonin (5-hydroxytryptamine [5-HT]) is a biogenic amine that plays numerous roles, through the regulation of systemic energy metabolism, in central and peripheral systems [7] as a neurotransmitter in the brain and as a hormone in peripheral tissues, respectively [8]. This review was conducted with the aim to compile the findings of recent studies of the relationship between 5-HT and hepatic metabolism and to discuss the role of 5-HT signaling as a potential treatment for NAFLD.
5-HT METABOLISM AND SIGNALING
L-tryptophan is an essential amino acid, and more than 90% of tryptophan is metabolized in the kynurenine pathways, and others are used to make 5-HT, melatonin, tryptamine, and indole-pyruvic acid [9]. 5-HT synthesis, both central and peripheral, is initiated through tryptophan hydroxylase (TPH)-mediated tryptophan hydroxylation that is closely related to tryptophan availability, kynurenine synthesis, and TPH, which is the rate-limiting enzyme [8] and occurs in two isoforms: TPH1 is mainly expressed in peripheral tissues and, centrally, TPH1 is mainly expressed in the pineal gland and at extremely low levels in the rest of the central nervous system (CNS) [10]; TPH2 is the predominant enzyme isoform for neuronal 5-HT synthesis in the CNS and the enteric nervous system (ENS) [7]. As 5-HT does not cross the blood–brain barrier, changes in activities of TPH1 and TPH1 alters 5-HT levels in peripheral tissues and CNS, respectively.
From tryptophan, TPH generates 5-hydroxytryptophan (5-HTP), which is then converted to 5-HT by aromatic acid decarboxylase (AADC). The 5-HT transporter (serotonin transporter [SERT]) facilitates 5-HT reuptake, and monoamine oxidase (MAO)-A catalyzes the oxidative deamination of 5-HT. Both SERT and MAO-A activities are important factors to determine 5-HT levels in the target organs [11]. Furthermore, 5-HT can be converted to N-acetyl-serotonin by arylalkylamine N-acetyltransferase and, subsequently, to melatonin by hydroxyindole O-methyltransferase in the pineal gland and retina [12]. Moreover, indoleamine 2,3-dioxygenase (IDO) metabolizes tryptophan, thereby allowing it to enter the kynurenine pathway, which accounts for approximately 95% of dietary tryptophan degradation [9].
Through receptor signaling pathways, 5-HT modulates various physiological and pathological processes. Most of the 5-HT-related biological processes are mediated through more than fourteen 5-HT receptors that are categorized into seven families [7], of which six are G-protein-coupled receptors and one is a ligand-gated cation channel (serotonin receptor 3 [HTR3]) (Table 1, Fig. 1) [7].
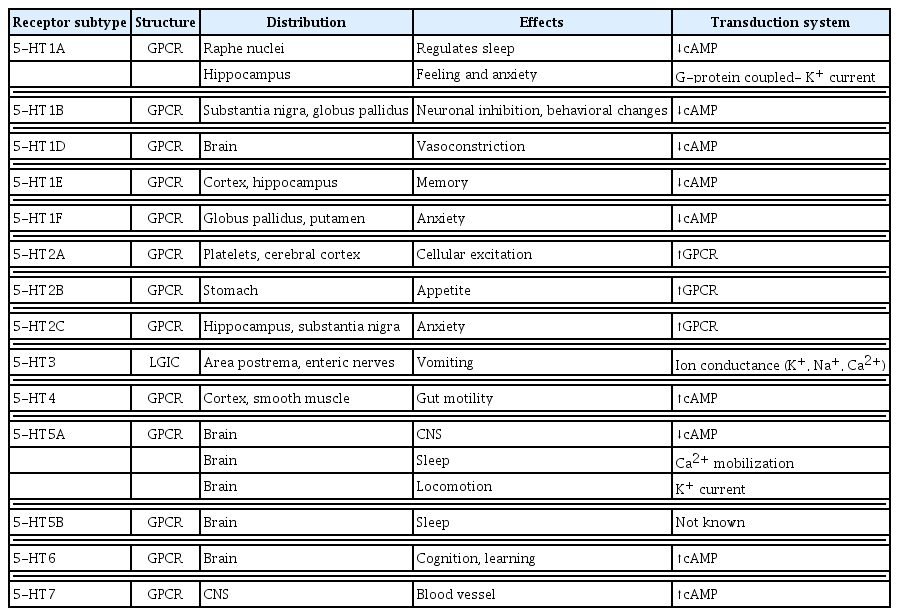
5-HT Receptors, Structure, Transduction System, and Tissue Expression

5-Hydroxytryptamine (5-HT) receptors and signaling pathways. The 5-HT1 and 5-HT5 receptors are Gi/Go-protein coupled receptors that inhibit adenylate cyclase (AC) and thus suppress the cyclic adenosine monophosphate (cAMP) downstream pathways. 5-HT2 receptors are Gq/G11-protein coupled receptors that activate phospholipase C, resulting in the activation of the inositol triphosphate (IP3) and diacylglycerol (DAG) downstream pathways. The only ligand-gated ion channel that can regulate membrane potential is the 5-HT3 receptor. The Gs-protein coupled receptors 5-HT4, 5-HT6, and 5-HT7 activate AC.
CENTRAL 5-HT AND ENERGY METABOLISM
Central 5-HT plays a major role in appetite control and systemic energy metabolism [13]. Several studies have elucidated the roles of 5-HT and its receptors in the CNS, and investigations in 5-HT receptor knockout (KO) mice have provided evidence that supports the role of central 5-HT in controlling appetite and/or bodyweight [14]. Via the 5-HT1B receptor, 5-HT stimulates the release of α-melanocyte-stimulating hormone in the proopiomelanocortin (POMC) neuron and suppresses the secretion of agouti-related protein (AgRP) in the orexigenic neuropeptide Y (NPY)/AgRP neuron [15], with a resultant decrease in appetite. The 5-HT2C receptor agonist lorcaserin (Belviq, Eisai Inc., Tokyo, Japan), which showed significant bodyweight reduction via appetite suppression, was approved in 2012 for the treatment of obesity by the Food and Drug Administration (FDA) [16] and saw widespread use as an anti-obesity medication until the FDA requested its withdrawal in 2020 due to an increased risk of cancer [17].
Central 5-HT signaling increases energy expenditure via the induction of thermogenic activity in brown adipose tissue (BAT) [14]. TPH2-positive neurons are a component of the neural circuitry between the brain and BAT [18]. The rostral raphé pallius, which regulates sympathetically mediated metabolism and the thermogenic activity of BAT, contains 5-HT neurons [19]. Microinjections of the 5-HT receptor antagonist, methysergide, into the intermediolateral cell column of the rat spinal cord suppressed cold-induced thermogenic activity in the BAT [19]. Central 5-HT deficiency resulted in the loss of thermoregulation and decreased the uncoupling protein 1 (UCP1) content in both BAT and inguinal white adipose tissue (WAT) [20]. Furthermore, through changes in the autonomic nervous system (ANS) and hormonal secretions, central 5-HT regulates peripheral glucose and lipid homeostasis [13], and POMC and AgRP neurons regulate glucose and lipid metabolism [13]. Thus, 5-HT2C receptor agonists regulate energy and glucose homeostasis and appetite via POMC neurons [21], and the AgRP neurons regulate hepatic glucose production [22].
PERIPHERAL 5-HT AND ENERGY METABOLISM
The majority (>95%) of 5-HT is synthesized by TPH1 in enterochromaffin cells of gut and stored in platelets [7]. The major peripheral organs, such as the heart, adipose tissue, pancreatic islets, and skeletal muscle, contain TPH1 and can synthesize 5-HT [23–26]. Furthermore, the ENS can produce 5-HT from TPH2 [27]. Thus, 5-HT can regulate the metabolic function of peripheral organs through autocrine/paracrine pathways.
Through 5-HT receptor transduction in the ENS, 5-HT regulates gut motility through the modulation of muscular peristaltic activity via motor and sensory functions. Submucosal and myenteric neurons that are involved in intestinal peristalsis, secretion, and sensation are regulated by 5-HT3 and 5-HT4 [28]. Ondansetron, a 5-HT3 receptor antagonist that is widely used to prevent nausea and vomiting, can induce side effects such as constipation and ileus. The 5-HT4 receptor accelerates propulsive motility and reduces visceral pain in the large intestine [29]. In vitro, a 5-HT4 receptor agonist can increase enteric neuronal development and survival [30].
Recent studies have suggested a possible relationship between gut microbiota and 5-HT signaling in the gastrointestinal tract. Compared to WT mice, Tph1-KO mice had different gut microbiota composition, which conferred a protective effect that resulted in a lower susceptibility to colitis [31], and which suggested that 5-HT regulates gut microbiota composition. In contrast, gut microbiota alters 5-HT levels in the colon and blood by directly regulating gastrointestinal tryptophan metabolism [32].
Pancreatic islets are important for glycemic control because they comprise α- and β-cells that secrete glucagon and insulin, respectively. During pregnancy, pancreatic β-cells synthesize 5-HT, which increases β-cell mass and glucose-stimulated insulin secretion via 5-HT2B and 5-HT3 receptor signaling, respectively [26,33]. In addition, 5-HT regulates insulin secretion in a diet-induced insulin resistance state [34] and controls adult β-cell mass by stimulating perinatal β-cell proliferation [35]. Furthermore, 5-HT regulates energy metabolism in adipose tissue. During fasting conditions, both lipolysis in adipose tissue and gluconeogenesis in the liver are increased. Sumara et al. [36] discovered that fasting increased the level of gut-derived 5-HT (GDS), which promoted lipolysis in WAT via 5-HT2b receptor signaling.
Tph1 and Tph2 double-KO mice, as well as Tph1-KO mice, have lower body weight [37,38]. In contrast, the body weights of gut-specific Tph1-KO mice are comparable to those of WT control mice [36]. Intriguingly, on a high-fat diet (HFD), adipose tissue-specific Tph1-KO mice had less weight gain than WT mice [24]. The HFD increased 5-HT levels in WAT with upregulated Tph1 expression, and 5-HT subsequently upregulated lipid accumulation in WAT via 5-HT2A receptor-induced lipogenesis [24,39]. These findings imply that 5-HT, in addition to GDS, possibly plays a role in the regulation of energy homeostasis, especially in WAT.
Furthermore, recent studies suggest possible roles of 5-HT in BAT. On an HFD, Tph1-KO mice showed increased energy expenditure when a peripheral TPH inhibitor (LP-533401) was used to confirm the obesogenic actions of peripheral 5-HT [24,40]. In adipocyte-specific Tph1-KO mice, 5-HT depletion induced Ucp1 and Dio2 expression in the BAT and subcutaneous WAT [24]. In brown fat, the 5-HT3 receptor is important for diet-induced thermogenesis, which significantly increased in Htr3a-KO (whole-body KO) mice fed an HFD [31]. Therefore, this lean phenotype could be attributed to either central or peripheral 5-HT effects. Future research is required to identify the primary 5-HT receptor that regulates BAT thermogenesis.
5-HT AND LIVER REGENERATION
The hepatocyte is unable to produce 5-HT; however, cholangiocytes and hepatic stellate cells (HSC) produce 5-HT in the liver [41]. Intrahepatic 5-HT-containing neurons and 5-HT receptors are distributed in the ANS, on branches of the hepatic artery, portal vein, bile duct, and the connective tissue of the interlobar septa [42]. Thus, neuronal signals from the ANS as well as circulating 5-HT from the gut regulates hepatic biology and metabolic functions, such as hepatic blood flow and regeneration.
In animal studies, 5-HT increased portal resistance in dogs [43] whereas the 5-HT2 receptor antagonist ketanserin reduced portal hypertension in rats [44]. The 5-HT2 receptor antagonist LY53857 reduced hepatic sinusoidal blood flow by acting on the HSC, which has both 5-HT2A and 5-HT2B receptors [45]. Furthermore, Kulinskii et al. [46,47] reported a major role of 5-HT in hepatic regeneration and wound healing using a murine partial hepatectomy (PHx) model. Tph1-KO mice failed to regenerate the liver after PHx, and 5-HT2 and 5-HT7 antagonists suppressed liver regeneration in post-PHx rats [48]. In a murine PHx model, the 5-HT2A receptor mediated liver regeneration [49]. The fibrogenic HSC have 5-HT2B receptor, and a 5-HT2B antagonist attenuated hepatic fibrosis and improved hepatic function in a murine chronic liver inflammation model [50]. Additionally, a recent study in a murine PHx model suggested a novel role of 5-HT, based on the activation the Hippo signaling pathway, in promoting hepatic regeneration [51].
Selective serotonin reuptake inhibitors (SSRIs) and serotonin noradrenaline reuptake inhibitors (SNRIs) reduced the intraplatelet 5-HT concentration. A recent clinical study reported that perioperative SSRI/SNRI intake was significantly associated with the high incidence of morbidity and postoperative hepatic dysfunction [52].
ROLE OF PERIPHERAL 5-HT IN NAFLD
The liver plays an important role in the regulation of both glucose and lipid homeostasis. To maintain plasma glucose levels during fasting, the liver increases glycogenolysis and gluconeogenesis. Sumara et al. [36] discovered that, via 5-HT2B receptor signaling during fasting, GDS promotes gluconeogenesis and inhibits hepatic glucose uptake. Furthermore, GDS inhibits intrahepatocyte glucose uptake via a glucose transporter 2-dependent mechanism.
In contrast, several studies found contradictory results. 5-HT boosted hepatic glucose uptake and increased intrahepatic fat content in a dog model [53]. In ob/ob mice, the duodenal 5-HT content increased, and treatment with a 5-HT3 receptor antagonist reduced the elevated 5-HT levels while increasing SERT activity in the duodenum. Moreover, the 5-HT3 receptor antagonist reduced fat content, inflammation, and necrosis in these mice [54]. Crane et al. [40] discovered an intrahepatic role for peripheral 5-HT in mice fed an HFD: WT mice became obese and developed fatty liver disease when fed an HFD, whereas Tph1-KO mice did not develop fatty liver on an HFD and had reduced hepatic fat accumulation.
Recently, Choi et al. [55] proposed 5-HT as a therapeutic target for NAFLD. Gut-specific Tph1-KO and liver-specific Htr2a-KO mice were both resistant to HFD-induced hepatic steatosis. In addition, the 5-HT2A antagonist sarpogrelate ameliorated hepatic steatosis in HFD-fed mice. Thus, GDS regulates hepatic lipid accumulation through 5-HT2A receptor signaling. Gene set enrichment analysis (GSEA) of the liver transcriptomes of HFD-fed liver-specific Htr2a-KO mice and WT littermates revealed that gene sets related to inflammation, fibrosis, and steatohepatitis were significantly downregulated in the liver of the former.
HSC are important cellular components of hepatic fibrosis and wound healing. 5-HT1A, 5-HT1F, 5-HT2A, 5-HT2B, and 5-HT7 receptors are expressed in rat and human HSC [56]. 5-HT works in tandem with platelet-derived growth factor to promote HSC proliferation [56]. In the diseased rat liver, 5-HT2B receptor was strongly associated with fibrotic tissue. 5-HT2 receptor antagonist treatment of HSC reduced their proliferation and increased their rate of apoptosis [56]. In human HSC lines, 5-HT2A receptor antagonists inhibited viability and wound healing [57]. Moreover, 5-HT2B receptor signaling activated HSC by increasing TGF-1 signaling [50]. Inhibition of the 5-HT2B receptor suppressed fibrogenesis and improved liver function in a murine carbon tetrachloride (CCl4)-induced cirrhosis model [50].
HUMAN MOLECULAR BIOLOGY AND CLINICAL EVIDENCE OF THE ASSOCIATION BETWEEN 5-HT AND NAFLD
Genetics and molecular research using human samples to identify underlying mechanisms and novel therapeutic targets for NAFLD have revealed potential roles for 5-HT signaling in NAFLD. The mRNA expressions of 5-HT2A and 5-HT2B receptor in human omental adipose tissue were significantly increased in obese subjects compared to lean subjects, and these gene expressions were positively correlated with serum aspartate aminotransferase and alanine aminotransferase levels [58]. A recent single-center clinical study found that levels of 5-hydroxyindoleacetic acid (5-HIAA; a metabolite of 5-HT) were significantly associated with hepatic events in NAFLD patients [59]. Compared to women with morbid obesity with normal liver histology, women with morbid obesity and NAFLD had decreased hepatic 5-HT2A and 5-HT2B receptor mRNA expressions [60]. Pantano et al. [61] recently used total RNA sequencing data to examine the molecular characteristics of histologically normal and NAFLD human livers, and suggested that the 5-HT2B receptor gene is one of the top 26 candidate genes related to the severity of liver fibrosis.
In clinical practice, many drugs, such as tricyclic antidepressant (TCAs), SSRIs, SNRIs, and 5-HT receptor agonists/antagonists, that regulate 5-HT signaling are widely used (Table 2). Clinical studies have reported significant associations between the abovementioned drug classes and liver disease. Patients who received TCAs (imipramine, tianeptine) [62,63] and SNRIs (venlafaxine, duloxetine) developed hepatic steatosis [64,65]. A clinical study showed that fluoxetine, an SSRI, increased serum triglyceride levels and hepatic lipid accumulation [66,67]. A clinical trial on the cardiometabolic benefits of lorcaserin, a 5-HT2C receptor agonist, showed that lorcaserin significantly improved fatty liver in obese adults, and the effect remained significant even after controlling for fat mass and change in body weight [68].

5-HT Receptor Agonists and Antagonists
CONCLUSIONS
The human body has two separate serotonergic systems: the central 5-HT and the peripheral 5-HT systems. The majority of research has concentrated on the mechanism whereby central 5-HT regulates mood and behavior. Increasingly, central 5-HT signaling has been used therapeutically for appetite suppression to facilitate weight loss. Central 5-HT regulates glucose and lipid metabolism as well as systemic energy metabolism by regulating the thermogenic activities of brown and beige fat [20,69,70]. Several recent studies have identified the role of peripheral 5-HT from gut enterochromaffin cells and other peripheral organs in the regulation of various biological functions and energy metabolism. Thus, increasing energy expenditure and improving insulin resistance by regulating peripheral 5-HT signaling have been proposed as a novel target for anti-obesity treatment [7,71]. 5-HT plays a major role in regulating hepatic circulation and regeneration, and peripheral 5-HT promotes fat deposition and lipogenesis. Moreover, 5-HT induces hepatic fibrosis and promotes HSC activation (Fig. 2). Inhibition of peripheral 5-HT signaling impaired the development of NAFLD in animal models [55,72]. In addition, clinical studies of drugs that affect 5-HT signaling have provided evidence of the pathophysiological importance of 5-HT in liver disease. Further molecular and clinical studies on the relationship between 5-HT and liver metabolism are needed to better understand the complex biology of 5-HT in the liver and to provide effective 5-HT- related therapeutic applications.
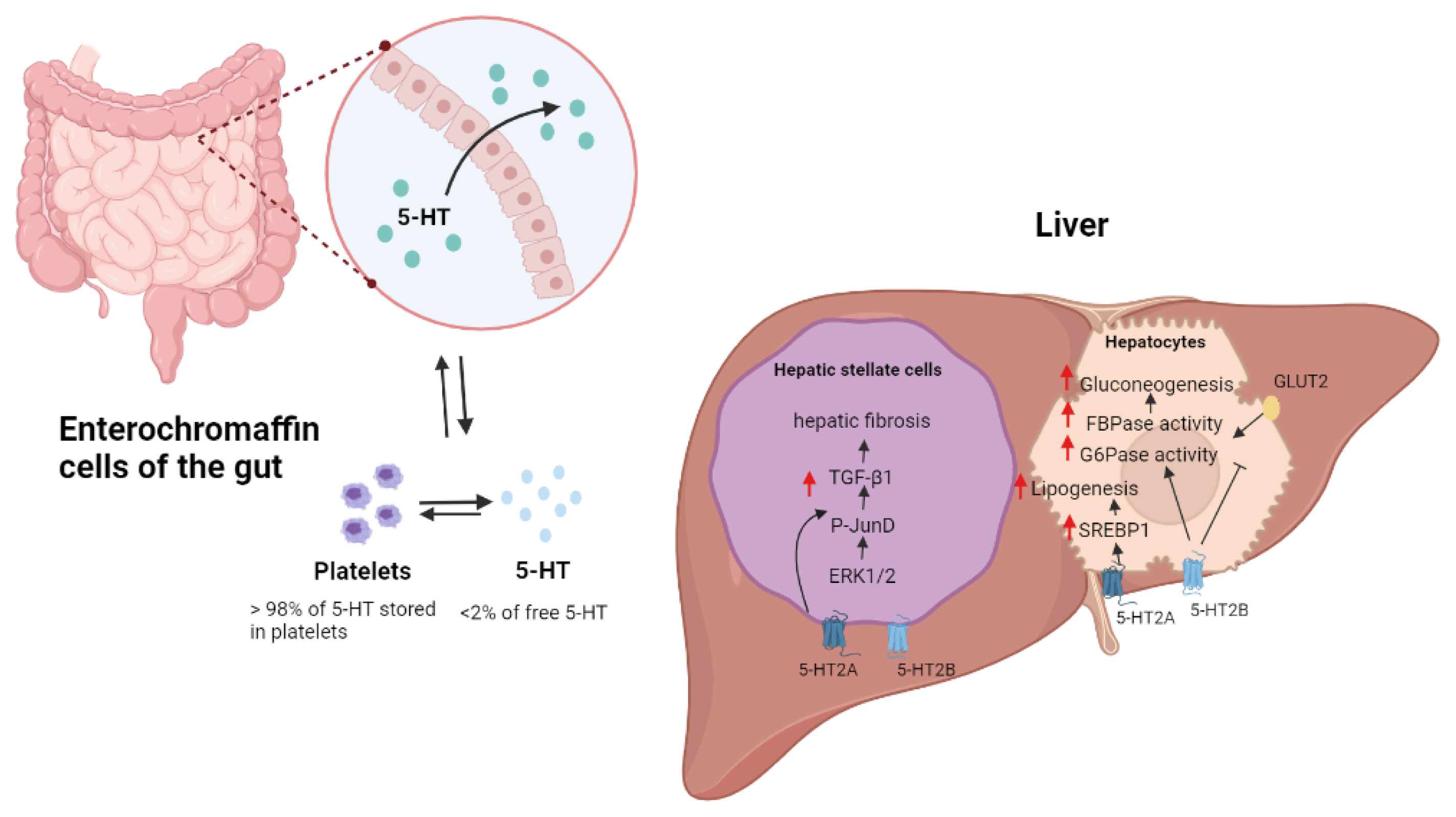
Metabolic functions of 5-hydroxytryptamine (5-HT) in liver. Most peripheral 5-HTs are derived from enterochromaffin cells of the gut. 5-HT regulates hepatic fibrosis in hepatic stellate cells (HSCs) by activating HSC-produced transforming growth factor (TGF)-signaling. Through the 5-HT2B receptor, 5-HT promotes gluconeogenesis in hepatocytes by increasing the activity of fructose 1,6-bisphosphatase (FBPase) and glucose 6-phosphatase (G6pase). 5-HT2B receptor signaling also inhibits glucose uptake by promoting the breakdown of glucose transporter 2 (GLUT2). The activation of the sterol-regulatory-element-binding protein 1 (SREBP1) signaling pathway by the 5-HT2A receptor increases lipogenesis in the liver. ERK1/2, extracellular signal–regulated protein kinase.
Acknowledgements
This research was supported by the Basic Science Research Program through the National Research Foundation of Korea (NRF), funded by the Ministry of Education (2020R1C1-C1004999) and KHIDI-AZ Diabetes Research Program.
Notes
CONFLICTS OF INTEREST
No potential conflict of interest relevant to this article was reported.