Articles
- Page Path
- HOME > Endocrinol Metab > Volume 29(2); 2014 > Article
-
Review ArticleRegulation of Adipocyte Differentiation via MicroRNAs
- You Hwa Son1*, Sojeong Ka2, A Young Kim2, Jae Bum Kim1,2
-
Endocrinology and Metabolism 2014;29(2):122-135.
DOI: https://doi.org/10.3803/EnM.2014.29.2.122
Published online: June 26, 2014
1Seoul National University School of Biological Sciences, Seoul National University, Seoul, Korea.
2Institute of Molecular Biology and Genetics, Seoul National University, Seoul, Korea.
- Corresponding author: Jae Bum Kim. School of Biological Sciences and Institute of Molecular Biology and Genetics, Seoul National University,1 Gwanak-ro, Gwanak-gu, Seoul 151-742, Korea. Tel: +82-2-880-5852, Fax: +82-2-878-5852, jaebkim@snu.ac.kr
- *Current affiliation: Division of Drug Discovery Research, Pharmacological Research Center, Korea Research Institute of Chemical Technology, Daejeon, Korea.
Copyright © 2014 Korean Endocrine Society
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
ABSTRACT
- Adipocyte differentiation, termed adipogenesis, is a complicated process in which pluripotent mesenchymal stem cells differentiate into mature adipocytes. The process of adipocyte differentiation is tightly regulated by a number of transcription factors, hormones and signaling pathway molecules. Recent studies have demonstrated that microRNAs, which belong to small noncoding RNA species, are also involved in adipocyte differentiation. In vivo and in vitro studies have revealed that various microRNAs affect adipogenesis by targeting several adipogenic transcription factors and key signaling molecules. In this review, we will summarize the roles of microRNAs in adipogenesis and their target genes associated with each stage of adipocyte differentiation.
- Adipose tissue plays key roles in energy storage, regulation of body temperature and absorption from mechanical collision. In a state of obesity, adipose tissue expands due to an increase in adipocyte size (hypertrophy) and/or number (hyperplasia), thereby inducing dysregulation of glucose and lipid metabolism. As a result, increased adipose tissue destroys whole body energy balance and enhances the risk of insulin resistance, hypertension, and dyslipidemia. Therefore, proper understanding of adipocyte differentiation would provide valuable information for designing comprehensive and effective therapeutic strategies against obesity.
- Adipocyte differentiation occurs in several stages, involves many signaling pathways, and progress depends on various stimuli such as nutrients and hormones. For example, adipogenesis is tightly controlled by a cascade of several transcription factors such as CCAAT/enhancer-binding proteins (C/EBPs) and peroxisome proliferator-activated receptor γ (PPARγ). Additionally, several signaling molecules, including wingless and INT-1 proteins (Wnts) and insulin, modulate adipogenesis. It has also been demonstrated that microRNAs are involved in adipocyte differentiation [1].
- MicroRNAs (miRNA) are 19 to 22 nucleotide fragments of noncoding RNA that play important roles in various cellular processes through posttranscriptional regulation of target genes. As many groups have reported that miRNAs actively participate in cell proliferation and differentiation [2,3,4], knowledge is constantly increasing related to miRNAs involvement in adipogenesis and fat metabolism. Although several reviews have recently been published on related topics [1,5,6], this review article will specifically describe the miRNA-mediated regulatory mechanism in adipocyte differentiation, particularly by updating and integrating the most recent information.
INTRODUCTION
- In vitro studies have suggested that the progression of adipocyte differentiation has, at least, two key steps: commitment and differentiation. Adipocytes are derived from pluripotent mesenchymal stem cells (MSCs) that have the capacity to develop into several cell types, including adipocytes and osteoblasts. Commitment or determination of MSCs' fate to differentiate into preadipocytes is caused by adipogenic differentiation signaling cues that have not yet been indentified [7]. Following this step, committed MSCs are specified for an adipogenic lineage and often lose their ability to differentiate into other cell lineages.
- In the differentiation step, committed preadipocytes derived from MSCs (e.g., 3T3-L1 cells) are differentiated into adipocytes after exposure to hormone cocktails such as insulin, dexamethasone and cyclic adenosine monophosphate (cAMP) activators [8]. Contact with these chemicals induces G1 phase-arrested 3T3-L1 cells to synchronously undergo, on average, two cycles of cell division, so called mitotic clonal expansion. During the cell cycle, clonal expansion is regulated by the Rb-E2F pathway, which is responsible for the G1-to-S transition. Rb inhibits the cell cycle by binding to, and repressing, the transcriptional activity of E2F. Upon hyperphosphorylation of Rb by cyclin-dependent kinases, E2F is released and promotes transcriptional activation of genes that encode cell-cycle regulators required for S phase entry; a process that initiates clonal expansion [9]. In growth-arrested preadipocytes, there are significant levels of Rb family p130 (pRB/p130). Inactivation of Rb2/p130 by phosphorylation enables clonal expansion. When cells exit the cell cycle, E2F loses its activity and terminal differentiation is initiated [10].
- Several signaling pathways highlight molecules such as bone morphogenic protein (BMP) and Wnt, which have been shown to be key molecules in the regulation of MSC commitment to adipocyte lineage and the differentiation of a subset of adipocytes. BMPs belong to the transforming growth factor β (TGF-β) family of growth factors, which consists of 14 family members. BMP-2 and BMP-4 have been implicated in adipogenesis and are thought to promote commitment of cells to adipogenic lineages [11,12,13,14]. The positive role of BMP-4 in adipocyte commitment has been demonstrated with several established cell lines. In C3H10T1/2 cells, exogenous BMP-4 activation induces potent adipocyte differentiation. In addition, a committed preadipocyte A33 cell line derived from C3H10T1/2 stem cells expresses and secretes BMP-4 at the same time point when exogenous BMP-4 is added to C3H10T1/2 cells for adipogenic differentiation. Furthermore, exposure of A33 cells to noggin, a naturally occurring BMP-4-binding antagonist, during this critical time window blocks subsequent differentiation [11]. The effect of BMP-2 is more complex. BMP-2 can enhance adipogenesis of C3H10T1/2 cells at low concentrations, but stimulates chondrocyte and osteoblast development at higher concentrations [14]. In preadipocytes, BMPs activate Sma and Mad related protein (Smad) signaling and regulate many target genes including cytoskeleton-associated proteins [12]. BMPs are also known as powerful cytokines that induce bone and cartilage formation. BMP-Smad signaling in this developmental context can activate runt-related transcription factor 2 (Runx2), osterix, Dlx5/6, and Sox9, which are essential transcription factors for osteogenesis and chondrogenesis [13].
- In addition to BMPs, the TGF-β superfamily member, TGF-β, is also involved in adipogenesis. In general, TGF-β signals through two types of transmembrane serine/threonine kinase receptors, type I and type II TGF-β receptors, and signaling effector Smads. Activation of Smad2 or Smad3 by TGF-β receptors results in heterodimerization with Smad4 and stimulates nuclear translocation of Smad complexes. In the nucleus, Smad proteins regulate transcription by binding to DNA and interacting with other transcription factors. During adipogenesis, TGF-β phosphorylates only Smad3, which then binds to C/EBPs and inhibits their transcriptional activity, including the ability to transactivate PPARγ [15]. Consistently, it has been demonstrated that TGF-β1 inhibits the early stages of 3T3-L1 differentiation [16] by promoting the proliferation of progenitor cells and hampering lipid accumulation [17]. Moreover, transgenic overexpression of TGF-β in adipose tissue inhibits differentiation in vivo [18].
- The Wnt family is made up of secreted glycoproteins that influence cell fate and development. Wnt proteins bind to frizzled receptors to stimulate signaling cascades through β-catenin-dependent (Wnt/β-catenin) and -independent pathways. The Wnt/β-catenin signaling pathway is often activated in preadipocytes and expression of Wnts declines after induction of differentiation [19,20]. It has been suggested that Wnt/β-catenin signaling enhances proliferation during commitment and mitotic clonal expansion [20]. Activated Wnt/β-catenin signaling enables the lymphoid-enhancer-binding factor/T-cell-specific transcription factor (LEF/TCF) family of transcription factors to activate Wnt target genes. In late stages of adipogenesis, Wnt/β-catenin signaling inhibits adipogenesis; however little is known about how Wnt inhibits adipogenesis through TCF/LEF [21].
- Many signaling pathways influence adipocyte differentiation. For instance, mitogen-activated protein kinase (MAPK) pathways may enhance adipogenesis through extracellular signal-regulated kinases (ERKs) and p38. Although it has been suggested that activation of ERK1 positively regulates adipogenesis during clonal expansion, activity has to be reduced after proliferation in order for adipogenesis to proceed [22]. Among various hormones, insulin plays a key role in adipogenesis. Insulin functions predominantly through insulin growth factor-1 receptor signaling, and the downstream signaling involves insulin receptor substrate (IRS), phosphoinositide 3-kinase (PI3K), PDK1, and AKT/protein kinase B (PKB) signaling. Insulin signaling can be transmitted to the adipogenic cascades in several different ways. IRS signaling is known to promote cAMP response element-binding (CREB) phosphorylation, thereby influencing adipogenesis [23]. Another way for insulin to deliver its signal is through inhibition of antiadipogenic forkhead box protein O (FOXO) transcription factors after AKT/PKB-mediated phosphorylation [24].
- Induction of adipogenesis by differentiation cocktails immediately phosphorylates and activates CREB, which in turn transcriptionally activates the C/EBPβ gene. Elevated C/EBPβ is somewhat inactive until it acquires DNA-binding activity through a series of phosphorylaton events by MAPK and GSK3β. Early activation of C/EBPβ, together with C/EBPδ, promotes gene expression of C/EBPα and PPARγ through C/EBP regulatory elements in their proximal promoters. C/EBPα induces adipogenic genes, and in vivo studies indicate an important role for this factor in the development of adipose tissue. PPARγ is the 'master regulator' of adipogenesis and is a necessary and sufficient factor for adipogenesis. Even though the C/EBPs are considered to be important transcription factors in adipogenesis, they cannot function efficiently without PPARγ [25]. PPARγ and C/EBPα coordinately control expression of a large group of genes that are required for adipocyte phenotypes. During adipogenesis, PPARγ and C/EBPα positively cross-activate each other through their respective C/EBP regulatory elements [25,26].
- Other key transcriptional factors are the Kruppel-like factors (KLFs). The KLFs are a large family of C2H2 zinc-finger proteins that regulate apoptosis, proliferation, and differentiation. This family of transcription factors contains both repressors and activators of transcription. Seven of the 17 known KLF proteins have been shown to be involved in different phases of adipocyte differentiation (i.e., KLF2 and 3 inhibit differentiation, whereas KLF4, 5, 6, 9, and 15 stimulate differentiation) [27].
MOLECULAR EVENTS DURING ADIPOCYTE DIFFERENTIATION
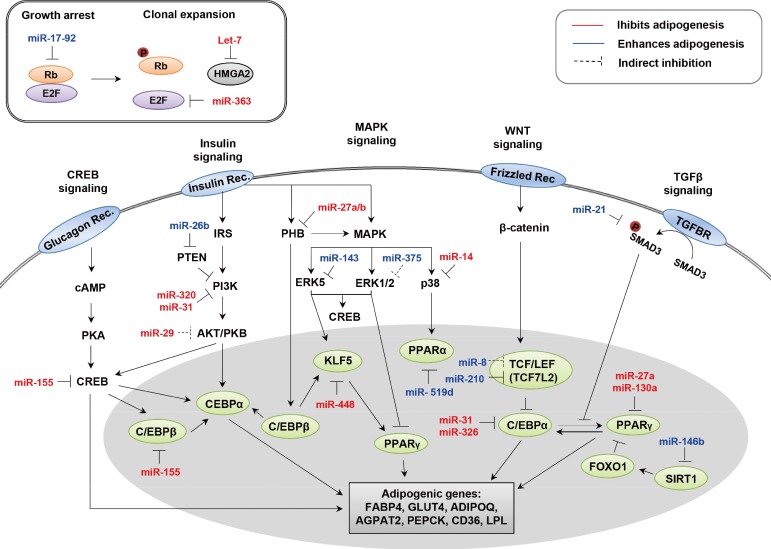
- A number of miRNAs and their targets have been implicated in adipogenesis. As summarized in Table 1 [1,2,28,29,30,31,32,33,34,35,36,37,38,39,40,41,42,43,44,45,46,47,48,49,50,51,52,53,54,55,56,57,58,59,60,61,62,63,64,65,66,67,68], some appear to enhance adipocyte differentiation while others inhibit adipogenesis in various model systems. An integrative model displaying the function of each miRNA in the progression of adipogenesis is depicted in Fig. 1.
ROLES OF miRNAs IN ADIPOGENESIS
- Adipocytes and osteoblasts originate from common MSCs. The differentiation of MSCs into adipocytes and osteoblasts is not only dependent on the mechanisms that determine a specific cell lineage, but also on the mechanisms that suppress the development of other lineages [69]. In mesenchymal progenitors, the selection between adipogenesis and osteogenesis is influenced by reciprocal regulation of different intracellular signals and transcription factors. Furthermore, previous studies have suggested that miRNAs are involved in both the lineage fate of MSCs and osteoblast differentiation [70].
- PPARγ regulates the whole process of adipogenesis including lineage commitment and differentiation. Thus, it appears that miRNAs specifically targeting PPARγ would also be modulators of adipogenesis. Several computational prediction programs have proposed that miR-27a and miR-130a may target PPARγ 3'-untranslated region (UTR) in a sequence-specific manner. In accordance with those predictions, it has been reported that both miR-27a and miR-130a indeed suppress adipocyte differentiation through PPARγ downregulation [28,29]. In 3T3-L1 cells, the levels of miR-27a and miR-130a are gradually decreased during adipogenesis, which is inversely correlated with expression levels of PPARγ. Furthermore, overexpression of miR-27a and miR-130a evidently suppresses adipocyte differentiation, concomitantly with PPARγ protein expression. These findings have suggested that both miR-27a and miR-130a negatively regulate PPARγ expression, leading to the repression of adipocyte differentiation. However, the roles of miR-27a and miR-130a in osteogenesis have not been elucidated. Thus, examining their roles in osteogenesis may provide insight into the physiological roles of miRNAs in the differentiation of MSCs to osteoblasts in bone tissue for therapeutic purposes.
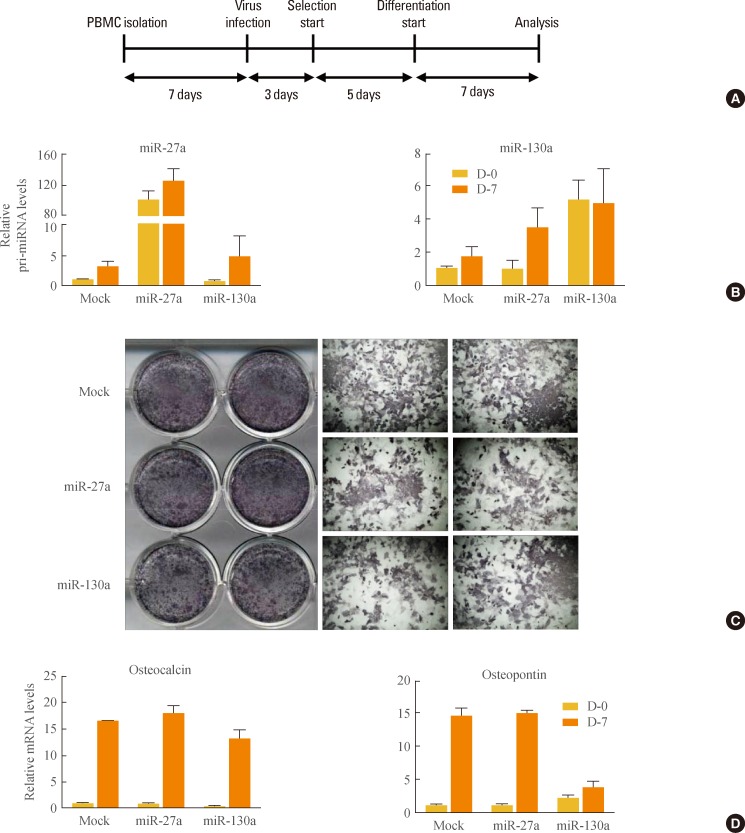
- To investigate whether miR-27a and miR-130a may regulate osteogenesis, isolated primary bone marrow cells (PBMCs) were retrovirally infected with miR-27a and miR-130a (Fig. 2A, B). After infection, PBMCs were maintained under osteogenic differentiation conditions. Alkaline phosphatase (ALP) staining showed that osteoblast differentiation was not influenced by miR-27a or miR-130a (Fig. 2C). Further, in PBMCs overexpressing miR-27a and miR-130a, mRNA expression levels of osteogenic marker genes such as osteopontin and osteocalcin were not systematically changed (Fig. 2D). These results reveal that miR-27a and miR-130a do not have significant effects on osteogenesis, implying that miR-27a and miR-130a may selectively affect adipogenesis, not osteogenesis.
- There are other known miRNAs that can control lineage determination. miR-124 is gradually upregulated during adipocyte differentiation of human bone marrow-derived mesenchymal stem cells (hMSCs). In 3T3-L1 cells, miR-124 overexpression promotes adipocyte differentiation via the suppression of Dlx5, accompanied by elevated fatty acid binding protein 4 (FABP4). Dlx5 is known as a pro-osteogenic transcription factor that determines cell fate in MSCs [71]. Thus, miR-124 has a proadipogenic effect by targeting Dlx5 [30]. Recent studies have demonstrated that miR-204, and its homolog miR-211, promote the induction of bone marrow stromal cell differentiation into adipocytes [31]. As shown by reporter assays, miR-211 directly targets 3'UTR of Runx2, which is a key transcription factor for osteoblast differentiation. Ectopic expression of miR-204 decreased Runx2 protein levels, whereas miR-204 knockdown significantly increased Runx2 protein levels. These data imply that miR-204 would suppress the action of Runx2 in osteoblast differentiation and enhance adipocyte differentiation.
LINEAGE DETERMINATION (COMMITMENT)
- Some miRNAs are involved in adipogenesis by regulating the RB-E2F pathway that controls mitotic clonal expansion. The miR-17-92 cluster has been reported to be upregulated during the clonal expansion stage of adipocyte differentiation, positively regulating adipogenesis [32]. Overexpression of miR-17-92 accelerates adipocyte differentiation and directly represses RB family Rb2/p130. On the contrary, miR-363 inhibits adipocyte differentiation by targeting E2F family member, E2F3 [72]. miR-363 overexpression downregulates expression levels of C/EBPα and PPARγ.
- In addition, let-7 negatively regulates adipogenesis by regulating the expression of high mobility group AT-hook2 (HMGA-2) [33]. Ectopic introduction of let-7 in 3T3-L1 and 3T3-F442A cells decreases mRNA levels of mature adipocyte markers such as FABP4 and PPARγ. Among several genes that have been previously shown to be targets of let-7, the most affected in 3T3-L1 is HMGA2, a transcription factor. Expression of HMGA2 is high during clonal expansion but turns off at terminal differentiation. HMGA2-deficient mice exhibit a reduction in adipose tissue [73], whereas overexpression in transgenic mice leads to an increase in fatty tissue [74]. This result depicts let-7 as an anti-adipogenic regulator that regulates the transition from clonal expansion to terminal differentiation.
miRNAs REGULATING MITOTIC CLONAL EXPANSION
- Some miRNAs control adipogenesis by blocking signal transduction pathways such as MAPK. miR-14 has a suppressive effect on fat metabolism by targeting p38 and MAPK in drosophila [34]. In flies, deletion of miR-14 increases the number of lipid droplets and the concentration of triacylglycerol. Similarly, miR-27a and miR-27b are involved in adipocyte differentiation. Computational prediction and luciferase assays have shown that miR-27a and miR-27b target prohibitin (PHB) [75], which is highly expressed in cells that rely heavily on mitochondrial function and has recently been implicated in adipogenesis [76]. In 3T3-L1 cells, overexpression of PHB inhibits insulin-induced adipogenesis, whereas in the absence of insulin, PHB facilitates adipogenesis through upregulation of MAPK/ERK signaling. PHB silencing by transfection with miR-27 impairs adipocyte differentiation in adipose tissue-derived stem cells (ASC). Likewise, protein levels of PHB, C/EBPβ, PPARγ, and aP2 are reduced in miR-27a- or miR-27b transfected ASCs. Moreover, ectopic expression of miR-27a or miR-27b attenuates lipid accumulation [35]. Considering all of these factors, miR-27a and miR-27b are therefore negative modulators of adipocyte differentiation by suppressing PHB.
- Many miRNAs have been reported to negatively regulate adipocyte differentiation by targeting insulin signaling and C/EBPs. In 3T3-L1 adipocytes, miR-320 and miR-29 decrease insulin activity by targeting PI3K directly [36] and AKT indirectly [37], respectively. However, it is unclear whether they can influence adipogenesis. Tang et al. [38] identified significant downregulation of miR-31 and miR-326 during adipogenesis of ADSCs. In silico prediction of target genes suggested that miR-31 may target phosphoinositide-3-kinase, class 2, alpha polypeptide (PIK3C2A) and C/EBPα, while miR-326 targets Ras association domain family member 1 (RASSF1) and AP2 associated kinase 1 (AAK1) [38]. In addition, expression of these predicted target genes is correlated with adipogenic differentiation. Another study has demonstrated that C/EBPα protein levels are downregulated by miR-31 expression in rats [39]. A recent report also demonstrated that miR-155 is an inhibitor of adipocyte differentiation. Ectopic expression of miR-155 decreased lipid accumulation as assessed by oil-red O staining in 3T3-L1 cells [40] and adipose tissue-derived preadipocytes [77]. Overexpression of miR-155 is associated with reduced protein levels of CREB and C/EBPβ, which are essential regulators during the early stages of terminal differentiation.
- Transcriptional or posttranscriptional processes related to adipogenesis are also controlled by many miRNAs. A bioinformatics approach has indicated that miR-448 targets KLF5 [41], and miR-448 is considered to be an inhibitor of adipocyte differentiation. Overexpression of miR-448 reduces KLF5 and inhibits adipocyte differentiation. mRNA levels of adipogenic marker genes such as C/EBPα and PPARγ, which are the targets of KLF5, are regulated by miR-448. In addition, miR-448 knockdown induces elevation of triglyceride concentration and upregulation of KLF5 expression, thereby promoting differentiation of 3T3-L1 preadipocytes. In 3T3-L1 cells, miR-224-5p can suppress adipocyte differentiation by targeting early growth response 2 (EGR2, also known as Krox20) [78]. EGR2 is a transcription factor that functions during early stages of adipocyte differentiation and enhances adipogenesis through C/EBPβ-dependent and -independent mechanisms. miR-224-5p is able to regulate fatty acid metabolism by directly targeting acyl-CoA synthetase long chain family member 4 (ACSL4). miR-138 has been implicated as an inhibitor of adipocyte differentiation through posttranscriptional regulation of EP300 interacting inhibitor of differentiation 1 (EID-1), which can promote adipocyte differentiation [42]. The role of miR-138 in human multipotent MSCs has been demonstrated by gain of function experiments. When miR-138 is overexpressed in hMSCs, lipid droplets are significantly reduced and the expression levels of C/EBPα and PPARγ decrease.
ANTIADIPOGENIC miRNAs
- It has been reported that several miRNAs may promote adipocyte differentiation by targeting MAPK signaling. In 3T3-L1 cells, expression of miR-143 increases during adipogenesis, and thereby stimulates adipocyte differentiation through inhibition of ERK5 via binding to the 3'-UTR of ERK5 mRNA [43]. Similarly, miR-375 overexpression increases the mRNA expression levels of C/EBPα, PPARγ, and adipocyte FABP (aP2) [36]. The number of lipid droplets is also increased by ectopic expression of miR-375 in 3T3-L1 cells. Furthermore, induction of miR-375 represses phosphorylation levels of ERK1/2. In 3T3-L1 cells, miR-375 knockdown elevates the phosphorylation level of ERK1/2 and decreases mRNA expression levels of C/EBPα, PPARγ, and aP2.
- Recent findings have suggested that miR-26b inhibits adipogenic differentiation in human preadipocytes [3]. Bioinformatics approaches and luciferase assays have revealed that the 3'UTR of the phosphatase and tensin homolog gene is a target of miR-26b. miR-26b overexpression enforces downregulation of adipogenic marker genes including aP2, C/EBPα and PPARγ. In addition, the abundance of lipid droplets and the triglyceride concentration are lowered in miR-26b-overexpressing cells. However, knockdown of miR-26b enhances adipocyte differentiation, accompanied with an increase in both adipocyte-specific marker genes and lipid droplets.
- Kennell et al. [44] have reported that miR-8 can inhibit the evolutionarily-conserved Wnt/Wingless pathway in drosophila. This regulation has been recapitulated with ST2 marrow stromal cells during adipogenesis. Overexpression of the miR-8 family (miR-200c-141 and miR-200b-200a-429 clusters) increases adipogenesis, FABP4, and lipid accumulation [44]. miR-8 indirectly targets the TCF transcription factor in Wnt signaling and directly targets CG32767, a zinc finger protein identified as a positive regulator of Wnt signaling in drosophila. Furthermore, miR-210 has been reported as a proadipogenic miRNA [79]. In 3T3-L1 cells, lentiviral overexpression of miR-210 enhances adipocyte differentiation, whereas inhibition of miR-210 suppresses expression of adipogenic marker genes. miR-210 enhances adipogenesis by inhibiting TCF7L2, which is a key transcription factor in Wnt signaling.
- According to the microarray results, the expression of miR-21 changes during adipocyte differentiation [80]. Oil-red O staining has demonstrated that miR-21 overexpression increases the abundance of lipid droplets and the size of adipocytes. In 3T3-L1 cells, miR-21 stimulates an increase in mRNA and protein levels of adiponectin. In addition, regulation of human ASC (hASC) differentiation by miR-21 is mediated by direct inhibition of TGF-β receptor 2 (TGFBR2) expression, which is closely related to the TGF-β signaling pathway. Recent findings suggest that miR-21 may enhance adipocyte differentiation of hASCs via inhibition of TGFBR2 [2]. miR-21 is known to have an adipogenic effect by targeting TGFBR2 in TGF-β signaling.
- In addition, miR-146b has been identified as a positive modulator of adipocyte differentiation via suppression of sirtuin 1 (SIRT1) [81]. SIRT1, a target of miR-146b, works as an inhibitor of adipocyte differentiation by inducing deacetylation of FOXO1. Expression patterns of miR-146b change significantly during adipocyte differentiation. In 3T3-L1 cells, ectopic expression of miR-146b increases expression of C/EBPα, PPARγ, and aP2. Furthermore, knockdown of miR-146b significantly decreases expression of adipogenic markers, body weight and visceral fat. Interestingly, miR-146b is highly expressed in the adipose tissue of obese mouse models such as DIO, diet-induced obesity, ob/ob, and db/db. This finding has implicated miR-146b as a positive regulator of accelerated adipocyte differentiation through modulation of SIRT1.
- It has also been shown that miR-519d is downregulated in obese individuals. In human subcutaneous adipose tissue, miR-519d overexpression results in an increase in adipocyte differentiation by targeting PPARα [45]. PPARα plays a crucial role in fatty acid oxidation in the liver, muscles and heart. Therefore, it is likely that miR-519d enhances adipogenesis by inhibiting fat burning through the downregulation of PPARα.
- Some miRNAs exhibit clear association with adipogenesis although their target genes or processes are unknown. For example, gain of function experiments have revealed that overexpression of miR-103 in preadipocytes upregulates expression of adipogenic genes such as PPARγ. Several miR-103 studies have been reported; however, there are inconsistencies between the miRNA profiles of mice and humans [46]. Thus, further research is necessary to delineate the role of miR-103 in adipocyte differentiation. miR-378 is also involved in adipocyte development, and is highly induced during adipocyte differentiation of ST2 cells [82]. Overexpression of miR-378 increases the size of lipid droplets and seems to promote the transcriptional activity of C/EBPα and C/EBPβ. Conversely, knockdown of miR-378 decreases the rate of triglyceride accumulation in ST2 cells. miR-132 has been identified as a biomarker of obesity.
- Numerous studies have revealed that miRNAs are associated with key metabolic parameters in obese patients. Microarray expression profiles have shown different circulating levels of miR-132 between obese and nonobese omental fat. Analysis of miR-132 has revealed that altered expression of miR-132 in blood is correlated with body mass index, fasting glucose, and glycosylated hemoglobin [6]. Among the targets of miR-132, CREB has been shown to be involved in regulation of appetite and glucose level. Thus, miR-132 expression may contribute to regulation of metabolic actions including energy homeostasis.
PROADIPOGENIC miRNAs
- Because obesity-related hyperplasia and hypertrophy are often associated with various metabolic disorders, detailed under standings of the molecular events regulating adipogenesis and lipid metabolism are important. Emerging evidence shows that miRNAs may directly or indirectly modulate adipocyte different iation [6]. The regulation of adipogenesis by miRNAs includes altering the expression of genes associated with each stage of adipocyte differentiation. In this regard, miRNAs can act on adipogenic factors in either positive or negative manners. Moreover, recent findings have proposed the idea that certain miRNAs could be biomarkers of obesity. However, many challenges remain. For example, a complex redundancy of miRNAs and target interactions may exist during the com plicated processes of adipogenesis, and the nature of these interactions has not been fully characterized. Therefore, further investigations are definitely required to elucidate the precise roles of these miRNAs and their regulatory mechanisms in adipogenesis. In conclusion, more research concerning the miRNAs involved in adipocyte differentiation would provide pathophysiological roles for these miRNAs and novel insight into obesity and its related metabolic diseases.
CONCLUSIONS
-
Acknowledgements
- This work was supported by the National Creative Research Initiative Program (2011-0018312), funded by the Ministry of Education, Science and Technology (MEST).
ACKNOWLEDGMENTS
- 1. McGregor RA, Choi MS. microRNAs in the regulation of adipogenesis and obesity. Curr Mol Med 2011;11:304–316. ArticlePubMedPMC
- 2. Kim YJ, Hwang SJ, Bae YC, Jung JS. MiR-21 regulates adipogenic differentiation through the modulation of TGF-beta signaling in mesenchymal stem cells derived from human adipose tissue. Stem Cells 2009;27:3093–3102. PubMed
- 3. Song G, Xu G, Ji C, Shi C, Shen Y, Chen L, Zhu L, Yang L, Zhao Y, Guo X. The role of microRNA-26b in human adipocyte differentiation and proliferation. Gene 2014;533:481–487. ArticlePubMed
- 4. Vimalraj S, Selvamurugan N. MicroRNAs: synthesis, gene regulation and osteoblast differentiation. Curr Issues Mol Biol 2012;15:7–18. PubMed
- 5. Alexander R, Lodish H, Sun L. MicroRNAs in adipogenesis and as therapeutic targets for obesity. Expert Opin Ther Targets 2011;15:623–636. ArticlePubMedPMC
- 6. Heneghan HM, Miller N, Kerin MJ. Role of microRNAs in obesity and the metabolic syndrome. Obes Rev 2010;11:354–361. ArticlePubMed
- 7. Tang QQ, Lane MD. Adipogenesis: from stem cell to adipocyte. Annu Rev Biochem 2012;81:715–736. ArticlePubMed
- 8. MacDougald OA, Lane MD. Transcriptional regulation of gene expression during adipocyte differentiation. Annu Rev Biochem 1995;64:345–373. ArticlePubMed
- 9. Burkhart DL, Sage J. Cellular mechanisms of tumour suppression by the retinoblastoma gene. Nat Rev Cancer 2008;8:671–682. ArticlePubMedPMCPDF
- 10. Fajas L, Debril MB, Auwerx J. Peroxisome proliferator-activated receptor-gamma: from adipogenesis to carcinogenesis. J Mol Endocrinol 2001;27:1–9. ArticlePubMed
- 11. Bowers RR, Kim JW, Otto TC, Lane MD. Stable stem cell commitment to the adipocyte lineage by inhibition of DNA methylation: role of the BMP-4 gene. Proc Natl Acad Sci U S A 2006;103:13022–13027. ArticlePubMedPMC
- 12. Huang HY, Hu LL, Song TJ, Li X, He Q, Sun X, Li YM, Lu HJ, Yang PY, Tang QQ. Involvement of cytoskeleton-associated proteins in the commitment of C3H10T1/2 pluripotent stem cells to adipocyte lineage induced by BMP2/4. Mol Cell Proteomics 2011;10:M110.002691.ArticlePubMed
- 13. Nishimura R, Hata K, Ikeda F, Ichida F, Shimoyama A, Matsubara T, Wada M, Amano K, Yoneda T. Signal transduction and transcriptional regulation during mesenchymal cell differentiation. J Bone Miner Metab 2008;26:203–212. ArticlePubMedPDF
- 14. Wang EA, Israel DI, Kelly S, Luxenberg DP. Bone morphogenetic protein-2 causes commitment and differentiation in C3H10T1/2 and 3T3 cells. Growth Factors 1993;9:57–71. ArticlePubMed
- 15. Choy L, Derynck R. Transforming growth factor-beta inhibits adipocyte differentiation by Smad3 interacting with CCAAT/enhancer-binding protein (C/EBP) and repressing C/EBP transactivation function. J Biol Chem 2003;278:9609–9619. ArticlePubMed
- 16. Zamani N, Brown CW. Emerging roles for the transforming growth factor-{beta} superfamily in regulating adiposity and energy expenditure. Endocr Rev 2011;32:387–403. ArticlePubMedPDF
- 17. Tsurutani Y, Fujimoto M, Takemoto M, Irisuna H, Koshizaka M, Onishi S, Ishikawa T, Mezawa M, He P, Honjo S, Maezawa Y, Saito Y, Yokote K. The roles of transforming growth factor-beta and Smad3 signaling in adipocyte differentiation and obesity. Biochem Biophys Res Commun 2011;407:68–73. ArticlePubMed
- 18. Clouthier DE, Comerford SA, Hammer RE. Hepatic fibrosis, glomerulosclerosis, and a lipodystrophy-like syndrome in PEPCK-TGF-beta1 transgenic mice. J Clin Invest 1997;100:2697–2713. ArticlePubMedPMC
- 19. Bennett CN, Ross SE, Longo KA, Bajnok L, Hemati N, Johnson KW, Harrison SD, MacDougald OA. Regulation of Wnt signaling during adipogenesis. J Biol Chem 2002;277:30998–31004. ArticlePubMed
- 20. Bowers RR, Lane MD. Wnt signaling and adipocyte lineage commitment. Cell Cycle 2008;7:1191–1196. ArticlePubMed
- 21. Singh R, Artaza JN, Taylor WE, Braga M, Yuan X, Gonzalez-Cadavid NF, Bhasin S. Testosterone inhibits adipogenic differentiation in 3T3-L1 cells: nuclear translocation of androgen receptor complex with beta-catenin and T-cell factor 4 may bypass canonical Wnt signaling to down-regulate adipogenic transcription factors. Endocrinology 2006;147:141–154. ArticlePubMedPDF
- 22. Bost F, Aouadi M, Caron L, Binetruy B. The role of MAPKs in adipocyte differentiation and obesity. Biochimie 2005;87:51–56. ArticlePubMed
- 23. Tseng YH, Butte AJ, Kokkotou E, Yechoor VK, Taniguchi CM, Kriauciunas KM, Cypess AM, Niinobe M, Yoshikawa K, Patti ME, Kahn CR. Prediction of preadipocyte differentiation by gene expression reveals role of insulin receptor substrates and necdin. Nat Cell Biol 2005;7:601–611. ArticlePubMedPDF
- 24. Nakae J, Kitamura T, Kitamura Y, Biggs WH 3rd, Arden KC, Accili D. The forkhead transcription factor Foxo1 regulates adipocyte differentiation. Dev Cell 2003;4:119–129. ArticlePubMed
- 25. Rosen ED, MacDougald OA. Adipocyte differentiation from the inside out. Nat Rev Mol Cell Biol 2006;7:885–896. ArticlePubMedPDF
- 26. Elberg G, Gimble JM, Tsai SY. Modulation of the murine peroxisome proliferator-activated receptor gamma 2 promoter activity by CCAAT/enhancer-binding proteins. J Biol Chem 2000;275:27815–27822. ArticlePubMed
- 27. Siersbaek R, Nielsen R, Mandrup S. Transcriptional networks and chromatin remodeling controlling adipogenesis. Trends Endocrinol Metab 2012;23:56–64. ArticlePubMed
- 28. Kim SY, Kim AY, Lee HW, Son YH, Lee GY, Lee JW, Lee YS, Kim JB. miR-27a is a negative regulator of adipocyte differentiation via suppressing PPARgamma expression. Biochem Biophys Res Commun 2010;392:323–328. ArticlePubMed
- 29. Lee EK, Lee MJ, Abdelmohsen K, Kim W, Kim MM, Srikantan S, Martindale JL, Hutchison ER, Kim HH, Marasa BS, Selimyan R, Egan JM, Smith SR, Fried SK, Gorospe M. miR-130 suppresses adipogenesis by inhibiting peroxisome proliferator-activated receptor gamma expression. Mol Cell Biol 2011;31:626–638. ArticlePubMed
- 30. Qadir AS, Woo KM, Ryoo HM, Baek JH. Insulin suppresses distal-less homeobox 5 expression through the up-regulation of microRNA-124 in 3T3-L1 cells. Exp Cell Res 2013;319:2125–2134. ArticlePubMed
- 31. Huang J, Zhao L, Xing L, Chen D. MicroRNA-204 regulates Runx2 protein expression and mesenchymal progenitor cell differentiation. Stem Cells 2010;28:357–364. ArticlePubMedPMC
- 32. Wang Q, Li YC, Wang J, Kong J, Qi Y, Quigg RJ, Li X. miR-17-92 cluster accelerates adipocyte differentiation by negatively regulating tumor-suppressor Rb2/p130. Proc Natl Acad Sci U S A 2008;105:2889–2894. ArticlePubMedPMC
- 33. Sun T, Fu M, Bookout AL, Kliewer SA, Mangelsdorf DJ. MicroRNA let-7 regulates 3T3-L1 adipogenesis. Mol Endocrinol 2009;23:925–931. ArticlePubMedPMCPDF
- 34. Xu P, Vernooy SY, Guo M, Hay BA. The Drosophila microRNA Mir-14 suppresses cell death and is required for normal fat metabolism. Curr Biol 2003;13:790–795. ArticlePubMed
- 35. Lin Q, Gao Z, Alarcon RM, Ye J, Yun Z. A role of miR-27 in the regulation of adipogenesis. FEBS J 2009;276:2348–2358. ArticlePubMedPMC
- 36. Ling HY, Wen GB, Feng SD, Tuo QH, Ou HS, Yao CH, Zhu BY, Gao ZP, Zhang L, Liao DF. MicroRNA-375 promotes 3T3-L1 adipocyte differentiation through modulation of extracellular signal-regulated kinase signalling. Clin Exp Pharmacol Physiol 2011;38:239–246. ArticlePubMedPMC
- 37. He A, Zhu L, Gupta N, Chang Y, Fang F. Overexpression of micro ribonucleic acid 29, highly up-regulated in diabetic rats, leads to insulin resistance in 3T3-L1 adipocytes. Mol Endocrinol 2007;21:2785–2794. ArticlePubMedPDF
- 38. Tang YF, Zhang Y, Li XY, Li C, Tian W, Liu L. Expression of miR-31, miR-125b-5p, and miR-326 in the adipogenic differentiation process of adipose-derived stem cells. OMICS 2009;13:331–336. ArticlePubMed
- 39. Sun F, Wang J, Pan Q, Yu Y, Zhang Y, Wan Y, Wang J, Li X, Hong A. Characterization of function and regulation of miR-24-1 and miR-31. Biochem Biophys Res Commun 2009;380:660–665. ArticlePubMed
- 40. Liu S, Yang Y, Wu J. TNFalpha-induced up-regulation of miR-155 inhibits adipogenesis by down-regulating early adipogenic transcription factors. Biochem Biophys Res Commun 2011;414:618–624. ArticlePubMed
- 41. Kinoshita M, Ono K, Horie T, Nagao K, Nishi H, Kuwabara Y, Takanabe-Mori R, Hasegawa K, Kita T, Kimura T. Regulation of adipocyte differentiation by activation of serotonin (5-HT) receptors 5-HT2AR and 5-HT2CR and involvement of microRNA-448-mediated repression of KLF5. Mol Endocrinol 2010;24:1978–1987. ArticlePubMedPMCPDF
- 42. Yang Z, Bian C, Zhou H, Huang S, Wang S, Liao L, Zhao RC. MicroRNA hsa-miR-138 inhibits adipogenic differentiation of human adipose tissue-derived mesenchymal stem cells through adenovirus EID-1. Stem Cells Dev 2011;20:259–267. ArticlePubMed
- 43. Esau C, Kang X, Peralta E, Hanson E, Marcusson EG, Ravichandran LV, Sun Y, Koo S, Perera RJ, Jain R, Dean NM, Freier SM, Bennett CF, Lollo B, Griffey R. MicroRNA-143 regulates adipocyte differentiation. J Biol Chem 2004;279:52361–52365. ArticlePubMed
- 44. Kennell JA, Gerin I, MacDougald OA, Cadigan KM. The microRNA miR-8 is a conserved negative regulator of Wnt signaling. Proc Natl Acad Sci U S A 2008;105:15417–15422. ArticlePubMedPMC
- 45. Martinelli R, Nardelli C, Pilone V, Buonomo T, Liguori R, Castano I, Buono P, Masone S, Persico G, Forestieri P, Pastore L, Sacchetti L. miR-519d overexpression is associated with human obesity. Obesity (Silver Spring) 2010;18:2170–2176. ArticlePubMed
- 46. Xie H, Lim B, Lodish HF. MicroRNAs induced during adipogenesis that accelerate fat cell development are downregulated in obesity. Diabetes 2009;58:1050–1057. ArticlePubMedPMC
- 47. Takanabe R, Ono K, Abe Y, Takaya T, Horie T, Wada H, Kita T, Satoh N, Shimatsu A, Hasegawa K. Up-regulated expression of microRNA-143 in association with obesity in adipose tissue of mice fed high-fat diet. Biochem Biophys Res Commun 2008;376:728–732. ArticlePubMed
- 48. Oskowitz AZ, Lu J, Penfornis P, Ylostalo J, McBride J, Flemington EK, Prockop DJ, Pochampally R. Human multipotent stromal cells from bone marrow and microRNA: regulation of differentiation and leukemia inhibitory factor expression. Proc Natl Acad Sci U S A 2008;105:18372–18377. ArticlePubMedPMC
- 49. Ling HY, Ou HS, Feng SD, Zhang XY, Tuo QH, Chen LX, Zhu BY, Gao ZP, Tang CK, Yin WD, Zhang L, Liao DF. CHANGES IN microRNA (miR) profile and effects of miR-320 in insulin-resistant 3T3-L1 adipocytes. Clin Exp Pharmacol Physiol 2009;36:e32–e39. ArticlePubMed
- 50. Jordan SD, Kruger M, Willmes DM, Redemann N, Wunderlich FT, Bronneke HS, Merkwirth C, Kashkar H, Olkkonen VM, Bottger T, Braun T, Seibler J, Bruning JC. Obesity-induced overexpression of miRNA-143 inhibits insulin-stimulated AKT activation and impairs glucose metabolism. Nat Cell Biol 2011;13:434–446. ArticlePubMedPDF
- 51. Yi C, Xie WD, Li F, Lv Q, He J, Wu J, Gu D, Xu N, Zhang Y. MiR-143 enhances adipogenic differentiation of 3T3-L1 cells through targeting the coding region of mouse pleiotrophin. FEBS Lett 2011;585:3303–3309. ArticlePubMed
- 52. El Ouaamari A, Baroukh N, Martens GA, Lebrun P, Pipeleers D, van Obberghen E. miR-375 targets 3'-phosphoinositide-dependent protein kinase-1 and regulates glucose-induced biological responses in pancreatic beta-cells. Diabetes 2008;57:2708–2717. ArticlePubMedPMC
- 53. Rajasethupathy P, Fiumara F, Sheridan R, Betel D, Puthanveettil SV, Russo JJ, Sander C, Tuschl T, Kandel E. Characterization of small RNAs in Aplysia reveals a role for miR-124 in constraining synaptic plasticity through CREB. Neuron 2009;63:803–817. ArticlePubMedPMC
- 54. Laine SK, Alm JJ, Virtanen SP, Aro HT, Laitala-Leinonen TK. MicroRNAs miR-96, miR-124, and miR-199a regulate gene expression in human bone marrow-derived mesenchymal stem cells. J Cell Biochem 2012;113:2687–2695. ArticlePubMed
- 55. Kloting N1, Berthold S, Kovacs P, Schon MR, Fasshauer M, Ruschke K, Stumvoll M, Bluher M. MicroRNA expression in human omental and subcutaneous adipose tissue. PLoS One 2009;4:e4699ArticlePubMedPMC
- 56. Qin L, Chen Y, Niu Y, Chen W, Wang Q, Xiao S, Li A, Xie Y, Li J, Zhao X, He Z, Mo D. A deep investigation into the adipogenesis mechanism: profile of microRNAs regulating adipogenesis by modulating the canonical Wnt/beta-catenin signaling pathway. BMC Genomics 2010;11:320ArticlePubMedPMC
- 57. Lee YS, Dutta A. The tumor suppressor microRNA let-7 represses the HMGA2 oncogene. Genes Dev 2007;21:1025–1030. ArticlePubMedPMC
- 58. Karbiener M, Fischer C, Nowitsch S, Opriessnig P, Papak C, Ailhaud G, Dani C, Amri EZ, Scheideler M. microRNA miR-27b impairs human adipocyte differentiation and targets PPARgamma. Biochem Biophys Res Commun 2009;390:247–251. ArticlePubMed
- 59. Wilfred BR, Wang WX, Nelson PT. Energizing miRNA research: a review of the role of miRNAs in lipid metabolism, with a prediction that miR-103/107 regulates human metabolic pathways. Mol Genet Metab 2007;91:209–217. ArticlePubMedPMC
- 60. Andersen DC, Jensen CH, Schneider M, Nossent AY, Eskildsen T, Hansen JL, Teisner B, Sheikh SP. MicroRNA-15a fine-tunes the level of Delta-like 1 homolog (DLK1) in proliferating 3T3-L1 preadipocytes. Exp Cell Res 2010;316:1681–1691. ArticlePubMed
- 61. Gu Z, Eleswarapu S, Jiang H. Identification and characterization of microRNAs from the bovine adipose tissue and mammary gland. FEBS Lett 2007;581:981–988. ArticlePubMed
- 62. Estep M, Armistead D, Hossain N, Elarainy H, Goodman Z, Baranova A, Chandhoke V, Younossi ZM. Differential expression of miRNAs in the visceral adipose tissue of patients with non-alcoholic fatty liver disease. Aliment Pharmacol Ther 2010;32:487–497. ArticlePubMed
- 63. Meerson A, Traurig M, Ossowski V, Fleming JM, Mullins M, Baier LJ. Human adipose microRNA-221 is upregulated in obesity and affects fat metabolism downstream of leptin and TNF-α. Diabetologia 2013;56:1971–1979. ArticlePubMedPMCPDF
- 64. Parra P, Serra F, Palou A. Expression of adipose microRNAs is sensitive to dietary conjugated linoleic acid treatment in mice. PLoS One 2010;5:e13005ArticlePubMedPMC
- 65. Zhu L, Chen L, Shi CM, Xu GF, Xu LL, Zhu LL, Guo XR, Ni Y, Cui Y, Ji C. MiR-335, an adipogenesis-related microRNA, is involved in adipose tissue inflammation. Cell Biochem Biophys 2014;68:283–290. ArticlePubMedPDF
- 66. Carrer M, Liu N, Grueter CE, Williams AH, Frisard MI, Hulver MW, Bassel-Duby R, Olson EN. Control of mitochondrial metabolism and systemic energy homeostasis by microRNAs 378 and 378*. Proc Natl Acad Sci U S A 2012;109:15330–15335. ArticlePubMedPMC
- 67. Jin W, Dodson MV, Moore SS, Basarab JA, Guan LL. Characterization of microRNA expression in bovine adipose tissues: a potential regulatory mechanism of subcutaneous adipose tissue development. BMC Mol Biol 2010;11:29ArticlePubMedPMC
- 68. Teleman AA, Maitra S, Cohen SM. Drosophila lacking microRNA miR-278 are defective in energy homeostasis. Genes Dev 2006;20:417–422. ArticlePubMedPMC
- 69. Muruganandan S, Roman AA, Sinal CJ. Adipocyte differentiation of bone marrow-derived mesenchymal stem cells: cross talk with the osteoblastogenic program. Cell Mol Life Sci 2009;66:236–253. ArticlePubMedPDF
- 70. Guo L, Zhao RC, Wu Y. The role of microRNAs in self-renewal and differentiation of mesenchymal stem cells. Exp Hematol 2011;39:608–616. ArticlePubMed
- 71. Holleville N, Mateos S, Bontoux M, Bollerot K, Monsoro-Burq AH. Dlx5 drives Runx2 expression and osteogenic differentiation in developing cranial suture mesenchyme. Dev Biol 2007;304:860–874. ArticlePubMed
- 72. Chen L, Cui J, Hou J, Long J, Li C, Liu L. A Novel Negative Regulator of Adipogenesis: MicroRNA-363. Stem Cells 2013.Article
- 73. Zhou X, Benson KF, Ashar HR, Chada K. Mutation responsible for the mouse pygmy phenotype in the developmentally regulated factor HMGI-C. Nature 1995;376:771–774. ArticlePubMedPDF
- 74. Arlotta P, Tai AK, Manfioletti G, Clifford C, Jay G, Ono SJ. Transgenic mice expressing a truncated form of the high mobility group I-C protein develop adiposity and an abnormally high prevalence of lipomas. J Biol Chem 2000;275:14394–14400. ArticlePubMed
- 75. Kang T, Lu W, Xu W, Anderson L, Bacanamwo M, Thompson W, Chen YE, Liu D. MicroRNA-27 (miR-27) targets prohibitin and impairs adipocyte differentiation and mitochondrial function in human adipose-derived stem cells. J Biol Chem 2013;288:34394–34402. ArticlePubMedPMC
- 76. Ande SR, Xu Z, Gu Y, Mishra S. Prohibitin has an important role in adipocyte differentiation. Int J Obes (Lond) 2012;36:1236–1244. ArticlePubMedPDF
- 77. Chen Y, Siegel F, Kipschull S, Haas B, Frohlich H, Meister G, Pfeifer A. miR-155 regulates differentiation of brown and beige adipocytes via a bistable circuit. Nat Commun 2013;4:1769ArticlePubMedPMCPDF
- 78. Peng Y, Xiang H, Chen C, Zheng R, Chai J, Peng J, Jiang S. MiR-224 impairs adipocyte early differentiation and regulates fatty acid metabolism. Int J Biochem Cell Biol 2013;45:1585–1593. ArticlePubMed
- 79. Liang WC, Wang Y, Wan DC, Yeung VS, Waye MM. Characterization of miR-210 in 3T3-L1 adipogenesis. J Cell Biochem 2013;114:2699–2707. ArticlePubMed
- 80. Kang M, Yan LM, Zhang WY, Li YM, Tang AZ, Ou HS. Role of microRNA-21 in regulating 3T3-L1 adipocyte differentiation and adiponectin expression. Mol Biol Rep 2013;40:5027–5034. ArticlePubMedPDF
- 81. Ahn J, Lee H, Jung CH, Jeon TI, Ha TY. MicroRNA-146b promotes adipogenesis by suppressing the SIRT1-FOXO1 cascade. EMBO Mol Med 2013;5:1602–1612. ArticlePubMedPMC
- 82. Gerin I, Bommer GT, McCoin CS, Sousa KM, Krishnan V, MacDougald OA. Roles for miRNA-378/378* in adipocyte gene expression and lipogenesis. Am J Physiol Endocrinol Metab 2010;299:E198–E206. ArticlePubMedPMC
References

MAPK, mitogen-activated protein kinase; ERK, extracellular signal-regulated kinase; MSC, mesenchymal stem cell; PI3K, phosphoinositide 3-kinase; T2D, type 2 diabetes; PTN, pleiotrophin; ORP, oxysterol-binding protein-related protein; PDK, phosphoinositide-dependent kinase; cAMP, cyclic adenosine monophosphate; PKA, protein kinase A; CREB, cAMP response element-binding; WNT, wingless and INT-1; TCF, T-cell-specific transcription factor; TGF-β, transforming growth factor β; TGFBR2, TGF-β receptor 2; hASC, human adipose tissue-derived stem cell; HMGA2, high mobility group AT-hook2; PPAR, peroxisome proliferator-activated receptor; hMADS, human multipotent adipose-derived stem; C/EBP, CCAAT/enhancer-binding protein; EID, EP300 interacting inhibitor of differentiation 1; KLF, Kruppel-like factor; Runx2, runt-related transcription factor 2; BMSC, bone marrow stromal cell; Smad1, Sma and Mad related protein 1; DLK1, delta-like homologue 1; O/E, overexpression.
Figure & Data
References
Citations
- A review of the role of transcription factors in regulating adipogenesis and lipogenesis in beef cattle
Belete Kuraz Abebe, Hongbao Wang, Anning Li, Linsen Zan
Journal of Animal Breeding and Genetics.2024; 141(3): 235. CrossRef - A review of the role of epigenetic studies for intramuscular fat deposition in beef cattle
Belete Kuraz Abebe, Jianfang Wang, Juntao Guo, Hongbao Wang, Anning Li, Linsen Zan
Gene.2024; 908: 148295. CrossRef - Transcriptome analysis of miRNAs during myoblasts adipogenic differentiation
Chengchuang Song, Xue Fang, Qi Wang, Yaqi Chen, Bei Zhao, Yanhong Wang, Xingtang Fang, Chunlei Zhang
Animal Biotechnology.2023; 34(4): 1406. CrossRef - Advances in the regulation of adipogenesis and lipid metabolism by exosomal ncRNAs and their role in related metabolic diseases
Cong Liu, Xilin Liu, Hong Li, Zhichen Kang
Frontiers in Cell and Developmental Biology.2023;[Epub] CrossRef - Indian gooseberry and barley sprout mixture prevents obesity by regulating adipogenesis, lipogenesis, and lipolysis in C57BL/6J mice with high-fat diet-induced obesity
Soo-Jeung Park, Jong-Lae Kim, Mi-Ryeong Park, Jong Wook Lee, Ok-Kyung Kim, Jeongmin Lee
Journal of Functional Foods.2022; 90: 104951. CrossRef - Interleukin-6 mimics insulin-dependent cellular distribution of some cytoskeletal proteins and Glut4 transporter without effect on glucose uptake in 3T3-L1 adipocytes
Maciej Błaszczyk, Małgorzata Gajewska, Marta Dymowska, Alicja Majewska, Tomasz Domoradzki, Adam Prostek, Rafał Pingwara, Magdalena Hulanicka, Katarzyna Grzelkowska-Kowalczyk
Histochemistry and Cell Biology.2022;[Epub] CrossRef - Heat-Killed Enterococcus faecalis Prevents Adipogenesis and High Fat Diet-Induced Obesity by Inhibition of Lipid Accumulation through Inhibiting C/EBP-α and PPAR-γ in the Insulin Signaling Pathway
Jin-Ho Lee, Keun-Jung Woo, Min-Ah Kim, Joonpyo Hong, Jihee Kim, Sun-Hong Kim, Kwon-Il Han, Masahiro Iwasa, Tack-Joong Kim
Nutrients.2022; 14(6): 1308. CrossRef - The delivery of miR-21a-5p by extracellular vesicles induces microglial polarization via the STAT3 pathway following hypoxia-ischemia in neonatal mice
Dan-Qing Xin, Yi-Jing Zhao, Ting-Ting Li, Hong-Fei Ke, Cheng-Cheng Gai, Xiao-Fan Guo, Wen-Qiang Chen, De-Xiang Liu, Zhen Wang
Neural Regeneration Research.2022; 17(10): 2238. CrossRef - Extracellular Vesicles from Adipose Tissue Could Promote Metabolic Adaptation through PI3K/Akt/mTOR
Jaime Delgadillo-Velázquez, Herminia Mendivil-Alvarado, Carlos Daniel Coronado-Alvarado, Humberto Astiazaran-Garcia
Cells.2022; 11(11): 1831. CrossRef - Adipocyte differentiation between obese and lean conditions depends on changes in miRNA expression
Yerim Heo, Hyunjung Kim, Jiwon Lim, Sun Shim Choi
Scientific Reports.2022;[Epub] CrossRef - Comparison of Selected Non-Coding RNAs and Gene Expression Profiles between Common Osteosarcoma Cell Lines
Mateusz Sikora, Katarzyna Krajewska, Klaudia Marcinkowska, Anna Raciborska, Rafał Jakub Wiglusz, Agnieszka Śmieszek
Cancers.2022; 14(18): 4533. CrossRef - Mesenchymal Stem Cell Secreted-Extracellular Vesicles are Involved in Chondrocyte Production and Reduce Adipogenesis during Stem Cell Differentiation
Yu-Chen Tsai, Tai-Shan Cheng, Hsiu-Jung Liao, Ming-Hsi Chuang, Hui-Ting Chen, Chun-Hung Chen, Kai-Ling Zhang, Chih-Hung Chang, Po-Cheng Lin, Chi-Ying F. Huang
Tissue Engineering and Regenerative Medicine.2022; 19(6): 1295. CrossRef - Tissue and circulating microRNAs as biomarkers of response to obesity treatment strategies
G. Catanzaro, T. Filardi, C. Sabato, A. Vacca, S. Migliaccio, S. Morano, E. Ferretti
Journal of Endocrinological Investigation.2021; 44(6): 1159. CrossRef - Benzyl Butyl Phthalate Induced Early lncRNA H19 Regulation in C3H10T1/2 Stem Cell Line
Jian Zhang, Mahua Choudhury
Chemical Research in Toxicology.2021; 34(1): 54. CrossRef - Inhibition of preadipocyte differentiation by Lycium barbarum polysaccharide treatment in 3T3-L1 cultures
Xiaochun Xu, Wenjuan Chen, Shukun Yu, Qian Lei, Lihong Han, Wenping Ma
Electronic Journal of Biotechnology.2021; 50: 53. CrossRef - MiR-25-3p regulates the differentiation of intramuscular preadipocytes in goat via targeting <i>KLF4</i>
Yu Du, Yue Zhao, Yong Wang, Qingyong Meng, Jiangjiang Zhu, Yaqiu Lin
Archives Animal Breeding.2021; 64(1): 17. CrossRef - MiR-208b Regulates Rabbit Preadipocyte Proliferation and Differentiation
Jiahao Shao, Ting Pan, Jie Wang, Tao Tang, Yanhong Li, Xianbo Jia, Songjia Lai
Genes.2021; 12(6): 890. CrossRef - Comparing the effect of cinnamaldehyde and metformin on expression of MiR320 and MiR26-b in insulin resistant 3T3L1 adipocytes
Yousof Naghiaee, Mahmood Vakili, Mohammad Mohammadi, Azra Mohiti, Javad Mohiti-Ardakani
Phytomedicine Plus.2021; 1(4): 100122. CrossRef - Lower miR‐26a levels in breastmilk affect gene expression in adipose tissue of offspring
Catalina A. Pomar, Francisca Serra, Andreu Palou, Juana Sánchez
The FASEB Journal.2021;[Epub] CrossRef - MicroRNA-378 regulates adipogenic differentiation in bovine intramuscular preadipocytes by targeting CaMKK2
Dongwei Li, Heng Wang, Yongmin Li, Changqing Qu, Yunhai Zhang, Hongyu Liu, Xiaorong Zhang
Adipocyte.2021; 10(1): 483. CrossRef - miR-6315 Attenuates Methotrexate Treatment-Induced Decreased Osteogenesis and Increased Adipogenesis Potentially through Modulating TGF-β/Smad2 Signalling
Ya-Li Zhang, Liang Liu, Yu-Wen Su, Cory J. Xian
Biomedicines.2021; 9(12): 1926. CrossRef - Inhibitory Effects of Pinostilbene on Adipogenesis in 3T3-L1 Adipocytes: A Study of Possible Mechanisms
You Chul Chung, Chang-Gu Hyun
International Journal of Molecular Sciences.2021; 22(24): 13446. CrossRef - Regulation of Methylase METTL3 on Fat Deposition
Gang Luo, Jialing Chen, Zhanjun Ren
Diabetes, Metabolic Syndrome and Obesity: Targets and Therapy.2021; Volume 14: 4843. CrossRef - miR-214-5p Regulating Differentiation of Intramuscular Preadipocytes in Goats via Targeting KLF12
Yu Du, Yong Wang, Yanyan Li, Quzhe Emu, Jiangjiang Zhu, Yaqiu Lin
Frontiers in Genetics.2021;[Epub] CrossRef - Screening and identification of MicroRNAs expressed in perirenal adipose tissue during rabbit growth
Guoze Wang, Guo Guo, Xueting Tian, Shenqiang Hu, Kun Du, Qinghai Zhang, Jingxin Mao, Xianbo Jia, Shiyi Chen, Jie Wang, Songjia Lai
Lipids in Health and Disease.2020;[Epub] CrossRef - Descending Expression of miR320 in Insulin-Resistant Adipocytes Treated with Ascending Concentrations of Metformin
Yousof Naghiaee, Reza Didehdar, Zahra Malekpour-Dehkordi, Fatemeh Pourrajab, Javad Mohiti-Ardakani
Biochemical Genetics.2020; 58(5): 661. CrossRef - Citrus aurantium L. Dry Extracts Ameliorate Adipocyte Differentiation of 3T3-L1 Cells Exposed to TNFα by Down-Regulating miR-155 Expression
Michele Campitelli, Antonella Desiderio, Giuseppe Cacace, Cecilia Nigro, Immacolata Prevenzano, Alessia Leone, Sonia de Simone, Pietro Campiglia, Pietro Formisano, Gregory A. Raciti, Francesco Beguinot, Claudia Miele
Nutrients.2020; 12(6): 1587. CrossRef - Metabolic Benefits of MicroRNA-22 Inhibition
Marc Thibonnier, Christine Esau
Nucleic Acid Therapeutics.2020; 30(2): 104. CrossRef - Differentially Expressed miRNA-Gene Targets Related to Intramuscular Fat in Musculus Longissimus Dorsi of Charolais × Holstein F2-Crossbred Bulls
Bilal Ahmad Mir, Henry Reyer, Katrin Komolka, Siriluck Ponsuksili, Christa Kühn, Steffen Maak
Genes.2020; 11(6): 700. CrossRef - MiR224-5p Inhibitor Restrains Neuronal Apoptosis by Targeting NR4A1 in the Oxygen-Glucose Deprivation (OGD) Model
Ling-Ling Liu, Shan Qiao, Mei-Ling Wang, Huai-Kuan Wu, Yong-Xin Su, Ke-Mo Wang, Xue-Wu Liu
Frontiers in Neuroscience.2020;[Epub] CrossRef - Adipogenesis: A Complex Interplay of Multiple Molecular Determinants and Pathways
Melvin A. Ambele, Priyanka Dhanraj, Rachel Giles, Michael S. Pepper
International Journal of Molecular Sciences.2020; 21(12): 4283. CrossRef - MicroRNA‑449a regulates the progression of brain aging by targeting SCN2B in SAMP8 mice
Ya‑Xin Tan, Ying Hong, Shui Jiang, Min‑Nan Lu, Shan Li, Bo Chen, Li Zhang, Tao Hu, Rui Mao, Rong Mei, Yan‑Bin Xiyang
International Journal of Molecular Medicine.2020;[Epub] CrossRef - Influence of adenovirus 36 seropositivity on the expression of adipogenic microRNAs in obese subjects
Víctor Manríquez, Alvaro Gutierrez, Alexis Morales, Roberto Brito, Monica Pavez, Jorge Sapunar, Luis Fonseca, Víctor Molina, Eugenia Ortiz, Maria Ines Barra, Camila Reimer, Maria Charles, Constance Schneider, Alvaro Cerda
International Journal of Obesity.2020; 44(11): 2303. CrossRef - Metformin downregulates miR223 expression in insulin-resistant 3T3L1 cells and human diabetic adipose tissue
Yousof Naghiaee, Reza Didehdar, Fatemeh Pourrajab, Masoud Rahmanian, Naeime Heiranizadeh, Azra Mohiti, Javad Mohiti-Ardakani
Endocrine.2020; 70(3): 498. CrossRef - Metabolic and energetic benefits of microRNA-22 inhibition
Marc Thibonnier, Christine Esau, Sujoy Ghosh, Edward Wargent, Claire Stocker
BMJ Open Diabetes Research & Care.2020; 8(1): e001478. CrossRef - Bta-miR-376a Targeting KLF15 Interferes with Adipogenesis Signaling Pathway to Promote Differentiation of Qinchuan Beef Cattle Preadipocytes
Xingyi Chen, Sayed Haidar Abbas Raza, Gong Cheng, Xinhao Ma, Jianfang Wang, Linsen Zan
Animals.2020; 10(12): 2362. CrossRef - The cross-talk between adipokines and miRNAs in health and obesity-mediated diseases
Ahmad Ghasemi, Seyed Isaac Hashemy, Mohsen Azimi-Nezhad, Alireza Dehghani, Jafar Saeidi, Mahnaz Mohtashami
Clinica Chimica Acta.2019; 499: 41. CrossRef - MicroRNAs and long noncoding RNAs: new regulators in cell fate determination of mesenchymal stem cells
Zixiang Wu, Shujing Liang, Wenyu Kuai, Lifang Hu, Airong Qian
RSC Advances.2019; 9(64): 37300. CrossRef - Harnessing adipogenesis to prevent obesity
Nida Haider, Louise Larose
Adipocyte.2019; 8(1): 98. CrossRef - MiR‐127 attenuates adipogenesis by targeting MAPK4 and HOXC6 in porcine adipocytes
Yun Gao, Yingqian Wang, Xiaochang Chen, Ying Peng, Fenfen Chen, Yulin He, Weijun Pang, Gongshe Yang, Taiyong Yu
Journal of Cellular Physiology.2019; 234(12): 21838. CrossRef - MicroRNA-425 controls lipogenesis and lipolysis in adipocytes
Renli Qi, Jing Wang, Qi Wang, Xiaoyu Qiu, Feiyun Yang, Zuohua Liu, Jinxiu Huang
Biochimica et Biophysica Acta (BBA) - Molecular and Cell Biology of Lipids.2019; 1864(5): 744. CrossRef - miRNA-7a-2-3p Inhibits Neuronal Apoptosis in Oxygen-Glucose Deprivation (OGD) Model
Zi-Bin Zhang, Ya-Xin Tan, Qiong Zhao, Liu-Lin Xiong, Jia Liu, Fei-Fei Xu, Yang Xu, Larisa Bobrovskaya, Xin-Fu Zhou, Ting-Hua Wang
Frontiers in Neuroscience.2019;[Epub] CrossRef - IL‐1α inhibits proliferation and adipogenic differentiation of human adipose‐derived mesenchymal stem cells through NF‐κB‐ and ERK1/2‐mediated proinflammatory cytokines
Xuerong Sun, Tangbin Zou, Changqing Zuo, Mingmeng Zhang, Benyan Shi, Zhiwen Jiang, Hongjing Cui, Xiaoxin Liao, Xiaoyi Li, Yuelian Tang, Yusheng Liu, Xinguang Liu
Cell Biology International.2018; 42(7): 794. CrossRef - MicroRNA‐224‐5p regulates adipocyte apoptosis induced by TNFα via controlling NF‐κB activation
Renli Qi, Jinxiu Huang, Qi Wang, Hong Liu, Ruisheng Wang, Jing Wang, Feiyun Yang
Journal of Cellular Physiology.2018; 233(2): 1236. CrossRef - Potential role of microRNAs in the regulation of adipocytes liposecretion and adipose tissue physiology
Giulia Maurizi, Lucia Babini, Lucio Della Guardia
Journal of Cellular Physiology.2018; 233(12): 9077. CrossRef - Chronic hyperinsulinemia induced miR-27b is linked to adipocyte insulin resistance by targeting insulin receptor
Ankita Srivastava, Kripa Shankar, Muheeb Beg, Sujith Rajan, Abhishek Gupta, Salil Varshney, Durgesh Kumar, Sanchita Gupta, Raj Kumar Mishra, Anil Nilkanth Gaikwad
Journal of Molecular Medicine.2018; 96(3-4): 315. CrossRef - Bta-miR-130a/b regulates preadipocyte differentiation by targeting PPARG and CYP2U1 in beef cattle
Xueyao Ma, Dawei Wei, Gong Cheng, Shijun Li, Li Wang, Yaning Wang, Xiaoyu Wang, Song Zhang, Hongbao Wang, Linsen Zan
Molecular and Cellular Probes.2018; 42: 10. CrossRef - Transdifferentiation of adipocytes to osteoblasts: potential for orthopaedic treatment
Daphne P L Lin, Crispin R Dass
Journal of Pharmacy and Pharmacology.2018; 70(3): 307. CrossRef - Characterization of miRNA transcriptome profiles related to breast muscle development and intramuscular fat deposition in chickens
Shouyi Fu, Yinli Zhao, Yuanfang Li, Guoxi Li, Yi Chen, Zhuanjian Li, Guirong Sun, Hong Li, Xiangtao Kang, Fengbin Yan
Journal of Cellular Biochemistry.2018; 119(8): 7063. CrossRef - Transcriptomic Analyses of Adipocyte Differentiation From Human Mesenchymal Stromal‐Cells (MSC)
Antonio Casado‐Díaz, Jaouad Anter, Sören Müller, Peter Winter, José Manuel Quesada‐Gómez, Gabriel Dorado
Journal of Cellular Physiology.2017; 232(4): 771. CrossRef - Downregulated miR-29a/b/c during Contact Inhibition Stage Promote 3T3-L1 Adipogenesis by Targeting DNMT3A
Yingjie Zhu, Guangyong Zheng, Huichao Wang, Yudong Jia, Ying Zhang, Yanfeng Tang, Wenlong Li, Yanan Fan, Xiaodong Zhang, Youwen Liu, Sanhong Liu, Makoto Kanzaki
PLOS ONE.2017; 12(1): e0170636. CrossRef - Potential miRNA involvement in the anti-adipogenic effect of resveratrol and its metabolites
Itziar Eseberri, Arrate Lasa, Jonatan Miranda, Ana Gracia, Maria P. Portillo, Cristina Óvilo
PLOS ONE.2017; 12(9): e0184875. CrossRef - Identification and characterization of differentially expressed miRNAs in subcutaneous adipose between Wagyu and Holstein cattle
Yuntao Guo, Xiuxiu Zhang, Wanlong Huang, Xiangyang Miao
Scientific Reports.2017;[Epub] CrossRef - Integrated analysis of mRNA and miRNA expression profiles in livers of Yimeng black pigs with extreme phenotypes for backfat thickness
Wentong Li, Yalan Yang, Ying Liu, Shuai Liu, Xiuxiu Li, Yingping Wang, Yanmin Zhang, Hui Tang, Rong Zhou, Kui Li
Oncotarget.2017; 8(70): 114787. CrossRef - Small non coding RNAs in adipocyte biology and obesity
Ez-Zoubir Amri, Marcel Scheideler
Molecular and Cellular Endocrinology.2017; 456: 87. CrossRef - Role of MicroRNA Regulation in Obesity-Associated Breast Cancer: Nutritional Perspectives
Ravi Kasiappan, Dheeran Rajarajan
Advances in Nutrition.2017; 8(6): 868. CrossRef - Biomolecular features of inflammation in obese rheumatoid arthritis patients: management considerations
Barbara Tolusso, Stefano Alivernini, Maria Rita Gigante, Gianfranco Ferraccioli, Elisa Gremese
Expert Review of Clinical Immunology.2016; 12(7): 751. CrossRef - Circulating microRNAs are deregulated in overweight/obese children: preliminary results of the I.Family study
Giuseppe Iacomino, Paola Russo, Ilaria Stillitano, Fabio Lauria, Pasquale Marena, Wolfgang Ahrens, Pasquale De Luca, Alfonso Siani
Genes & Nutrition.2016;[Epub] CrossRef - Role of MicroRNAs in NAFLD/NASH
Gyongyi Szabo, Timea Csak
Digestive Diseases and Sciences.2016; 61(5): 1314. CrossRef - MicroRNA‐29b promotes the adipogenic differentiation of human adipose tissue‐derived stromal cells
Xi‐Mei Zhang, Li‐Hong Wang, Dong‐Ju Su, Dan Zhu, Qiu‐Ming Li, Mei‐Hua Chi
Obesity.2016; 24(5): 1097. CrossRef - MiR-181a-5p regulates 3T3-L1 cell adipogenesis by targeting <italic>Smad7</italic> and <italic>Tcf7l2</italic>
Dan Ouyang, Lifeng Xu, Lihua Zhang, Dongguang Guo, Xiaotong Tan, Xiaofang Yu, Junjie Qi, Yaqiong Ye, Qihong Liu, Yongjiang Ma, Yugu Li
Acta Biochimica et Biophysica Sinica.2016; 48(11): 1034. CrossRef - Metformin-suppressed differentiation of human visceral preadipocytes: Involvement of microRNAs
Koji Fujita, Hisakazu Iwama, Kyoko Oura, Tomoko Tadokoro, Kayo Hirose, Miwako Watanabe, Teppei Sakamoto, Akiko Katsura, Shima Mimura, Takako Nomura, Joji Tani, Hisaaki Miyoshi, Asahiro Morishita, Hirohito Yoneyama, Keiichi Okano, Yasuyuki Suzuki, Takashi
International Journal of Molecular Medicine.2016; 38(4): 1135. CrossRef - Systematic study of cis-antisense miRNAs in animal species reveals miR-3661 to target PPP2CA in human cells
Jian Wang, Zongcheng Li, Bailong Liu, Guangnan Chen, Ningsheng Shao, Xiaomin Ying, Ya Wang
RNA.2016; 22(1): 87. CrossRef - Oxidative stress, redox regulation and diseases of cellular differentiation
Zhi-Wei Ye, Jie Zhang, Danyelle M. Townsend, Kenneth D. Tew
Biochimica et Biophysica Acta (BBA) - General Subjects.2015; 1850(8): 1607. CrossRef - Noncoding RNAs, cytokines, and inflammation-related diseases
José Luiz Marques-Rocha, Mirian Samblas, Fermin I. Milagro, Josefina Bressan, J. Alfredo Martínez, Amelia Marti
The FASEB Journal.2015; 29(9): 3595. CrossRef - Polymorphism in miR-31 and miR-584 binding site in the angiotensinogen gene differentially influences body fat distribution in both sexes
Jan Machal, Jan Novak, Renata Hezova, Filip Zlamal, Anna Vasku, Ondrej Slaby, Julie Bienertova-Vasku
Genes & Nutrition.2015;[Epub] CrossRef - Mitochondria-related miR-141-3p contributes to mitochondrial dysfunction in HFD-induced obesity by inhibiting PTEN
Juan Ji, Yufeng Qin, Jing Ren, Chuncheng Lu, Rong Wang, Xiuliang Dai, Ran Zhou, Zhenyao Huang, Miaofei Xu, Minjian Chen, Wei Wu, Ling Song, Hongbing Shen, Zhibin Hu, Dengshun Miao, Yankai Xia, Xinru Wang
Scientific Reports.2015;[Epub] CrossRef - MicroRNAs in Bone Balance and Osteoporosis
Junying Chen, Min Qiu, Ce Dou, Zhen Cao, Shiwu Dong
Drug Development Research.2015; 76(5): 235. CrossRef - Articles in 'Endocrinology and Metabolism' in 2014
Won-Young Lee
Endocrinology and Metabolism.2015; 30(1): 47. CrossRef - MicroRNA regulatory networks in human adipose tissue and obesity
Peter Arner, Agné Kulyté
Nature Reviews Endocrinology.2015; 11(5): 276. CrossRef - Epigenomics, gestational programming and risk of metabolic syndrome
M Desai, J K Jellyman, M G Ross
International Journal of Obesity.2015; 39(4): 633. CrossRef - Expression Profiling and Structural Characterization of MicroRNAs in Adipose Tissues of Hibernating Ground Squirrels
Cheng-Wei Wu, Kyle K. Biggar, Kenneth B. Storey
Genomics, Proteomics & Bioinformatics.2014; 12(6): 284. CrossRef
 PubReader
PubReader Cite
Cite